UWO
-- Neuroscience Graduate Program
Neuroscience 9500: Notes
for J. A. Kiernan's lecture.
For comfortable reading,
adjust the width of your web browser's window to about two-thirds of
the width of the screen.
This two-hour lecture has two themes.
There are
downloadable printer-friendly PDF files for both themes.
- 1. The
methods used to acquire knowledge of the ways in which different parts
of the nervous system are functionally interconnected.
(Click here for shortcut.)
- 2. The
somatic sensory pathways as an example of related systems of
interconnected neurons.
(Click here for shortcut.)
Where there
is a link,
it is usually to a picture or diagram. Click on the link to view the
illustration. Other links are to files that you can download.
Neuroanatomical
tracing
methods
The
file neurmeth.pdf
(click to download) contains somewhat longer accounts of
neuroanatomical methods and somatosensory pathways than this web page.
You need Adobe
Acrobat or the free Adobe Acrobat Reader program to read this file,
which is a shortened version of the accounts of these subjects in Barr's The Human Nervous System
(9th edition, 2008) [ WL101.B268h.2009
in the UWO Sciences Library ].
Traditional
approaches have provided most of our knowledge of
connectivity in the human brain and spinal cord. These include:
- Clinical consequeces of lesions
that destroy groups of neuronal cell bodies or sever fibre tracts
- Staining anterograde degeneration
in sections of the brain, post mortem.
- Includes Marchi
method (degenerating myelin black> and
- stains for normal
myelin (negative in degenerated tracts).
- Electrical stimulation, especially
of sites in the cerebral cortex in conscious patients undergoing
neurosurgical procedures under local anaesthesia
|
Modern
approaches have generally confirmed the
traditional body of knowledge of human neuroanatomy.
- Imaging techniques such as
computerized tomography (CT) and nuclear magnetic resonance imaging
(MRI) show extent of lesions at the same time that they cause
disability, allowing more accurate clinicopathological correlation than
is possible with post mortem examination.
- Functional imaging is based on
changes in blood flow, oxygen usage or electrical activity, and can
show parts of the brain that are relatively more metabolically active
than others.
Techniques include PET, fMRI, electroencephalography and
magnetoencephalography.
|
Experiments
with animals are needed to provide
detailed neuroanatomical information such as exact cells of origin and
sites of termination of axons.
- Degeneration methods include
silver stains that are selective for early degenerative changes in
axons (rather than myelin). These are seldom used today.
- Axonally transported tracers can
be
- Retrograde: Taken
up by synaptic terminals and moved to the cell body. Examples are
horseradish peroxidase, peroxidase- or fluorochrome-labelled
lectins, and several simple fluorescent dyes. Some viruses
such as pseudorabies, are retrogradely transported. They propagate in
the neuron, are transferred to its presynaptic terminals and then
transported to the cell bodies of the presynaptic neurons.
- Anterograde: Taken
up by cell bodies and dendrites and transported along axons to synaptic
terminals.
Various lectins are used this way, and there are fluorochromes that can
be microinjected into individual neuronal cell bodies
|
Somatic
sensory pathways.
The
file basicneuro.pdf (click
to download) is a 35-page summary of the anatomy and functional
connectivity of the human central nervous system. Somatic sensory
pathways are reviewed on pages 23-25. The pathways are also reviewed in
the second half of the file neurmeth.pdf. You need Adobe
Acrobat or the free Adobe Acrobat Reader program to read these files.
Neuronal
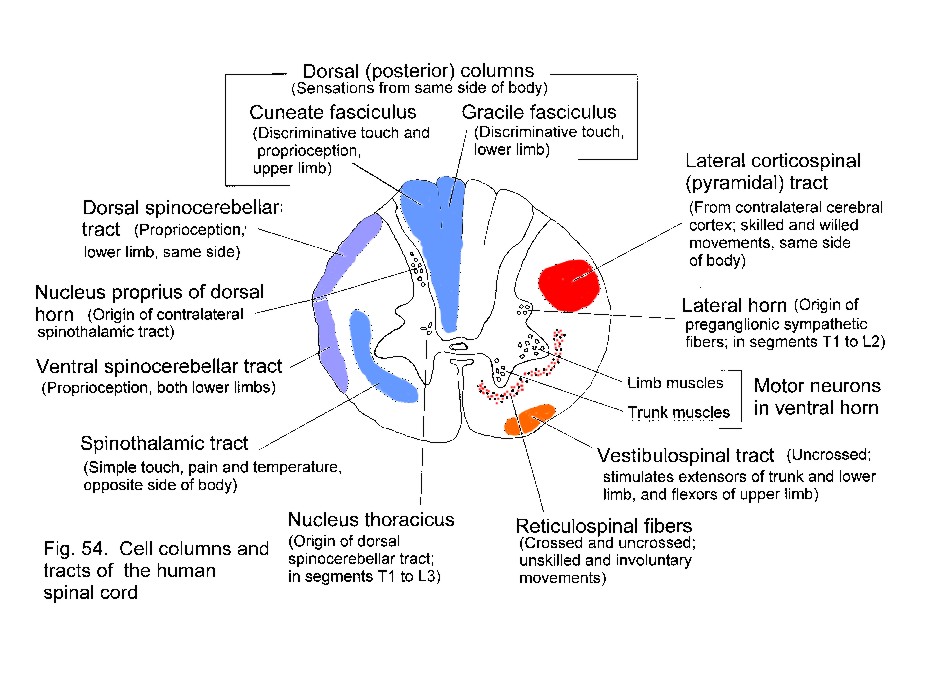
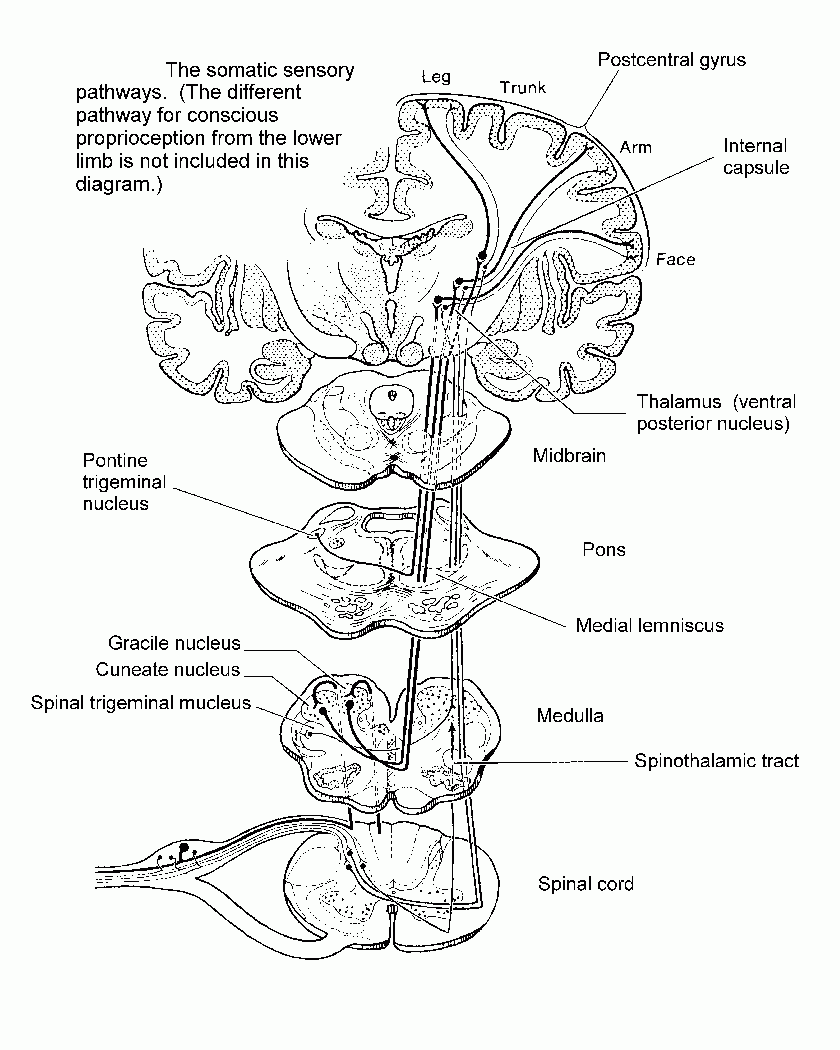
signals from skin and deeper structures are segregated in the spinal
cord. Transmission to the thalamus and cerebral cortex may occur
through the spinothalamic tract or through the dorsal funiculi and
medial lemniscus.
For pain,
temperature and the less discriminative aspects of touch,
neurons in the dorsal horn have axons that cross in the spinal cord and
ascend as the spinothalamic tract, which is
laterally situated in the spinal cord and brain stem.
For discriminative
touch and for conscious proprioception, the axons of
primary sensory neurons ascend ipsilaterally in the dorsal
funiculus
(either gracile or cuneate fasciculus)
and end in the gracile or cuneate
nucleus. Fibers arising in these nuclei cross in the
medulla and ascend
in the medial lemniscus, which is near the midline
in the medulla and
shifts to a lateral location in the midbrain.
For conscious proprioception from the lower
limb there is an additional pathway,
involving the dorsal spinocerebellar tract and nucleus Z in the
medulla.
Consequently a lesion
that transects the gracile fasciculus in the cervical spinal cord does
not cause complete loss of position sense in the lower limbs.
Proprioceptive sensation is lost if the lesion is larger, extending
ventrally and laterally into the territory of the dorsal
spinocerebellar tract.
| Reminder -
Positions of tracts in spinal cord
(Use
your web browser's Back button (right mouse click
in Netscape) to return from the picture to this web page.)
|
Click here to
download sompaths.exe (1266393 bytes).
This is a free-standing executable slide show. (It needs MS
Windows 95 or later. Download the file to a directory on your hard
drive. Run it by typing sompaths, either at the Run prompt
(shown when you click the Start
icon at the bottom of the screen) or at a command prompt (found in the
Accessories folder). |
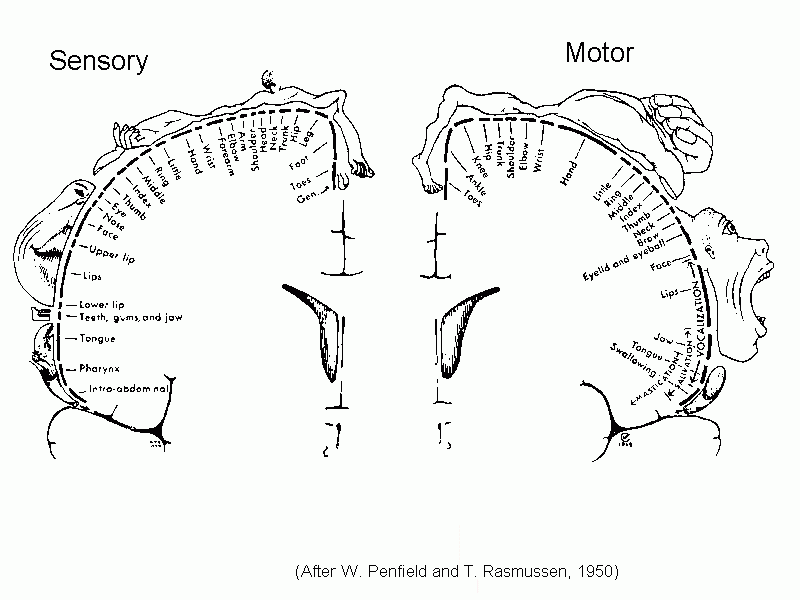
Both the
spinothalamic tract and the medial lemniscus end in the VPL nucleus of the thalamus.
This thalamic nucleus projects to the primary somesthetic
cortex of the postcentral gyrus, where
the
contralateral half of the body is represented as an upside-down
homunculus.
Diagram summarizing the somatic sensory
pathways
The somatic
sensory pathways for the head involve the trigeminal
sensory
nuclei and
their projections to the contralateral VPM
thalamic nucleus.
Primary afferent fibers for touch end in
the pontine trigeminal nucleus.
Pain and temperature fibers descend in the spinal trigeminal tract
before ending in
the caudal part of its nucleus. This pathway will be
discussed later, in connection with
the central connections of cranial nerve V.
Lesions
in the spinal cord and brain stem can affect the
somesthetic pathways
separately, causing dissociated sensory loss.
| What
sensory loss results from a destructive in the
lateral part of the medulla, involving the left spinothalamic tract and
sensory
nuclei of the left trigeminal nerve (but sparing other somatic sensory
pathways)?
Click
here for the answer (but
think first!)
|
 |
The main
pathways are supplemented by others, especially for pain, which
involve the reticular formation and thalamic nuclei other than the VPL
or VPM .
The cerebral
cortex is necessary for localizing the source of a painful stimulus and
for
the recognition of objects by touch.
A
note about nuclei in sensory pathways.
The synapses between the main neurons in a
pathway are not simply "relays," which would serve
no useful purpose. Through connections from other parts of the CNS, and
with the involvement
of local interneurons, the incoming signals are modified for onward
transmission.
For example:
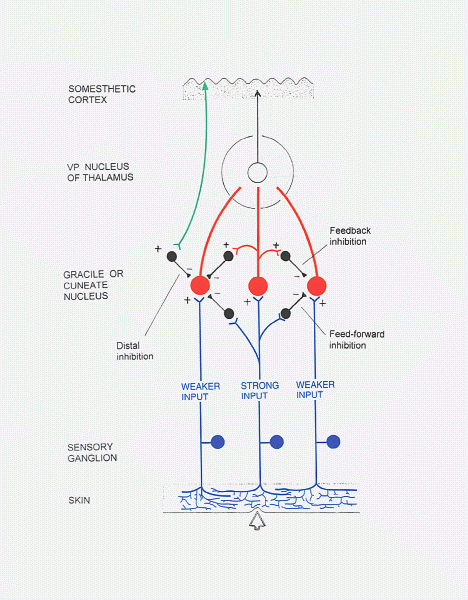
| In the gracile and cuneate nuclei, lateral inhibition
(feed-forward and feedback types) sharpens the perception of the most
strongly
stimulated part of a receptive field. There is also remote
inhibition
by corticobulbar fibres in these nuclei. |
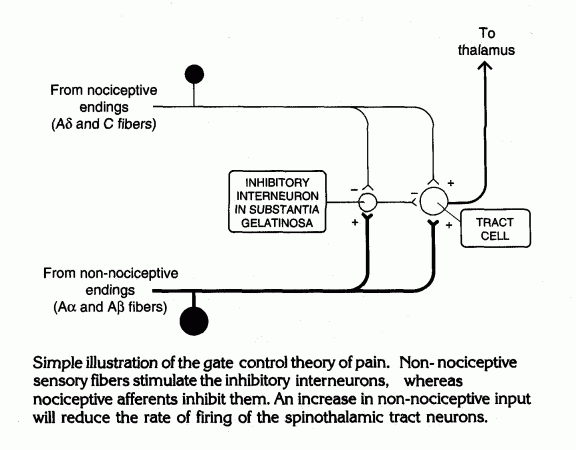
In the dorsal horn of the spinal cord, large and small
diameter axons contact
inhibitory interneurons and tract cells that send pain signals to the
thalamus. Balance between the two types of input constitutes the gate control mechanism
for pain. |
| Pain
is a big subject! It is mentioned only briefly
in the lecture and in these notes. A direct pathway from certain dorsal
horn neurons (Waldeyer cells)
to the mediodorsal (MD) thalamic nucleus is also involved. One of
several targets of neurons in MD is the anterior part of the cingulate
gyrus, on the medial surface
of the cerebral hemisphere. The anterior cingulate cortex recognizes
pain as a nasty sensation caused by injury or disease. Surgical lesions
are sometimes made in
the anterior part of the cingulate gyrus to relieve suffering that
cannot be relieved in any other way. After such surgery the patient
still feels the pain but
does not find the sensation intolerable. |
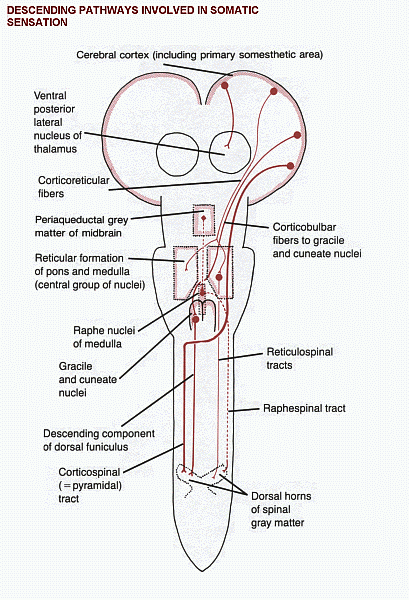
Descending
pathways that influence sensation
The proper
interpretation of the outside world and the state of the body itself
would be
impossible if every impulse in every sensory axon were to be brought
eventually to the
cerebral cortex. An editing system is necessary, so that the cortex can
select the sensory
information worthy of conscious attention while leaving more humble
duties to the
spinal cord, brain stem, and cerebellum. The editing function is
carried out by descending
fibres that terminate in the sites of origin of the ascending tracts.
Some of these
descending pathways are shown in
a diagram (Click here).
Descending
tracts modify activity in the ascending systems at three levels:
1. In
the dorsal horn of the spinal gray matter. This is
the site of termination of large
numbers of corticospinal fibers, mostly from the postcentral gyrus.
Other axons
ending in the dorsal horn come from the reticular formation and from
the gracile
and cuneate nuclei. One of the reticulospinal projections, the
raphespinal tract,
is notable for inhibiting the upward transmission of signals concerned
with
pain. It originates in the raphe nuclei, in the midline of the medulla,
and the unmyelinated serotonergic
axons of the tract are lateral to the tip of the dorsal horn.
The raphe nuclei are themselves stimulated by neurons in the
periaqueductal
gray matter of the midbrain.
Electrical stimulation
of the periaqueductal gray
causes prompt relief of pain and has occasionally been done clinically
for this
purpose. A curious observation is that stimulation for a few minutes
can produce
analgesia lasting several hours.
2. In
the brain stem. Large numbers of fibers descend
from the somatosensory
area of the cerebral cortex to the gracile and cuneate nuclei. They are
presumed to influence the
medial lemniscus system. Corticobulbar fibers also end in the
trigeminal sensory nuclei.
The suffix "-bulbar"
refers to the termination
of axons in the brain stem. The term "corticonuclear"
is sometimes used for axons of cortical neurons that end in
nuclei of cranial nerves. However, many descending fibers
that influence cranial nerves end near, but not within, the nuclei.
The less precise term "corticobulbar" is therefore preferred.
3. In
the thalamus. The VPL and VPM nuclei project to the
first somatosensory area of
the cerebral cortex. These thalamic nuclei also receive input from the
same cortical
areas.
(This isn't the only
thalamocortical
projection that is reciprocated; they all are.)
Recommended
reading
You should
read the appropriate pieces in the textbook for Neuroscience 9500:
Kandel, Schwarz & Jessell: Principles of Neural Science,
4th ed. [UWO Sciences Library: WL102.P9547
2000]. Neuroanatomical tracing
methods are covered in more than one chapter.
Somatic sensory pathways receive rather brief treatment, but
that's adequate for this course.
= = = = = = = = = = =
If you want
to learn more than you need to know for the exam, I will be happy to
recommend additional readings. My office is Rm 453 in the Medical
Sciences Building at UWO. Do drop in.
I can always
(= almost
always) take
a break to advise students about Neuroanatomy or Histochemistry. For a
wide range of free instructional resources and research-related items
follow the links from my personal UWO web page at http://publish.uwo.ca/~jkiernan or
look me up with Google. Notes for my current neuroanatomical
courses can be found by entering neuroanatomy uwo in
a Google search box.
John A.
Kiernan
Dept of Anatomy & Cell Biology
University of Western Ontario
London, Canada.
________________________________
Last updated 3rd November 2008
1. Contralateral
loss of pain and temperature sensation below
the head.
2. Ipsilateral loss of
pain and temperature sensation in the head (face, mouth,
pharynx etc).
3. Discriminative
touch and proprioception are not impaired. This lesion cause
other effects too (Wallenberg's syndrome).
Click here
to return to Somatic sensation
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}