
(Small plan of the nervous system) (Big picture of the whole human nervous system)
ANNOUNCEMENTS APPEAR HERE.
(http://www.uwo.ca/anatomy/grad/531a/531a.html). |
This web site contains introductory remarks and notes relating toHuman Neuroanatomy lectures by J.A.Kiernan.
Larger, better quality versions of the graphics are available by clicking on nearby links. Use your browser's Back button to return from the picture to this document.
|
J. A. Kiernan MB, ChB, PhD, DSc
Professor, Department of Anatomy & Cell Biology
Room 453, Medical Sciences Building, U.W.O.
If you have questions about the subject matter of this course, you are
always welcome to ask them at any reasonable time. My office is Room 453, Medical
Sciences Building. You do not need to make an appointment. Call 661-2111 ext 86822 if you want to check that
I'm there or ask a question on the phone. If you attend in person you will get better service. Talking on the phone is
helpful only for highly specific questions that have short answers.
You may also ask for very brief answers to questions by email: kiernan[AT]uwo.ca
Please come and see me as soon as you have difficulty understanding a topic.
Do not wait until the week immediately before an exam!
Instructional resources. Ask Mr Michael Wu of the
Department of Anatomy and Cell Biology (Room 440) if you wish to study in the
Anatomy Museum on the 4th floor (Dent. Sci. Bldg. Room 4002). Our museum contains many
neuroanatomy specimens and models.
For the lab classes you will need a
CD-ROM, specially prepared for use in conjunction with this course
by Mr Michael Wu. Unfortunately there is a charge of $5 to cover the cost
of production of the CD-ROM.
Dr D. G. Montemurro, formerly of the Department
of Anatomy at UWO, has produced numerous VHS videotapes that explain
the anatomy of the nervous system as it is seen in practical classes. Some
will be shown in 350a classes. All are available in the Taylor Library. Several of these tapes include more details than are needed for Anatomy 350a.
The required level of detail is that presented in the lectures and shown
on the CD-ROM.
For students who like such things, two sets of interactive
Sample exam questions are available for downloading. There is also a
sample of the video lab exam
that will show you what to expect in the practical part of the mid-term tests.
You can also download a
glossary of neuroanatomical terms.
Instant neuroanatomy (BasicNeuro.pdf). For an illustrated summary of the subject,
click here. The file BasicNeuro.pdf can be
read on screen or printed. (Use an inkjet printer if possible; most of
the illustrations are in colour.)
(You need the free
Adobe Acrobat Reader
to read or print PDF files. If you don't have this program, click on the link to download it from adobe.com)
If you understand everything in
BasicNeuro.pdf
and are familiar with the contents of the lab CD-ROM, you will certainly obtain at least a B grade for the course.
The plan. The first two lectures
introduce the major parts of the adult nervous system and explain how
they develop in the embryo. Two lectures on the peripheral nervous
system follow.
The next five weeks are devoted to
the anatomy of the central nervous system. There are one-hour lectures,
but the anatomy is studied principally by examination of whole and
dissected specimens of human brains in the Wednesday lab classes.
The mid-term test has two parts: multiple choice questions based on the lectures, and a video practical exam based on the lab classes.
Note! It is not possible to
separate the lecture material completely from the lab material because
many structures and functions are covered in both types of class.
For a sample of the type of questions in the video practical exam,
click here
to download the file SampleVP.exe. Run the file (needs Windows 95 or later) to see a sample "lab" exam with five questions.
The remainder of the course
consists of lectures on systemic or functional neuroanatomy. For these,
the teachers assume that the students remember most of the
topographical neuroanatomy from the first half of the course. Not all
the functional systems of the brain are included in this course. The
last lecture brings together the pathways and the topographical
neuroanatomy by showing that localized damage at different sites can
produce a wide variety of functional deficits. Much of the material in
this last lecture will be revision.
The final examination will
cover the whole subject, but most of the questions will related to
material covered in the second half of the course. Questions in both
exams will be set by the people who give the corresponding lectures.
The topics listed below correspond to the lecture titles.
Click on a link , below, for notes on the topic.
Within each topic there are clickable links that show text notes, diagrams, downloadable
files etc. Use your web browser's "back" button to return from a link to this page.
Asterisks (***)
indicate lectures by Dr W. Rushlow, who provides paper handouts. These
links are not active in this version of the web notes for Anatomy 350a.
(They are available, however, for graduate students taking the course
as 530a.)
Return to beginning of Anatomy 530a home page
Introduction to Anatomy 350/530. Major parts of the central and peripheral nervous systems.
Now for the neuroanatomy! Let's get started.
The CNS.
The central nervous system (CNS) is a tube, closed at both ends. The thickness of the wall of the tube varies, being greatest at the rostral (cranial) end. The complicated shape of the brain is due partly to the need to fold it up to fit into the skull. The CNS is made of fragile, gelatinous tissue. It is contained entirely within the axial skeleton (cranium and vertebral column).
These are the major divisions of the CNS:
In this list the most commonly used names are in bold type.
Spinal cord.
[Cavity: central canal]
Rhombencephalon or hindbrain
(consisting of medulla, pons and
[Cavities: central canal & 4th ventricle]
cerebellum).
[No internal cavity but near 4th ventricle]
Mesencephalon or midbrain.
[Cavity: cerebral aqueduct]
Prosencephalon or forebrain (consisting of the
diencephalon, and the telencephalon
[Cavity: 3rd ventricle]
or cerebral hemispheres, each of these divisions
being composed of many smaller parts.
[Cavities: lateral ventricles]
Nerves connect the CNS with all other parts of the body. A nerve is ensheathed in collagenous connective
tissue and can withstand the changes in pressure and tension associated with moving jointed limbs. Nerves go out
from the axial skeleton by way of foramina [foramen
= hole] in the cranium (cranial nerves) and
between vertebrae (spinal nerves). This general plan applies to all
vertebrate animals - even ones that don't have limbs or bony
skeletons.
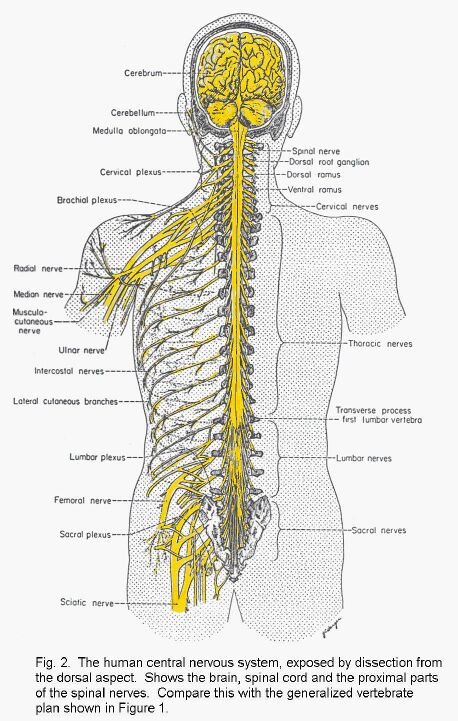
(Small plan of the nervous system)
(Big picture of the whole human nervous system)
Strategy for coping with terminology.
The greatest difficulty confronting the beginning student of neuroanatomy is becoming
familiar with a large number of names of parts. Use the index or the glossary of your textbook to look up
each new word you encounter. Do not attempt to learn the definition unless the word either
(a) applies to structure associated with a function that you have been told about. An example is the
calcarine sulcus, associated with vision.
(b) is something conspicuous enough to be a landmark relating to the positions of other structures. For example,
the lateral sulcus is near to areas of the cerebral cortex concerned with hearing, understanding what's heard, formulation of
speech, the sense of taste, and other parts of the forebrain with known functions and connections.
The PNS.
The peripheral nervous system (PNS) is composed of nerves and ganglia. A ganglion is a collection of neuronal cell bodies outside the CNS. Typically a ganglion is a lump on a nerve, but many of the ganglia associated with internal organs are of microscopic size.
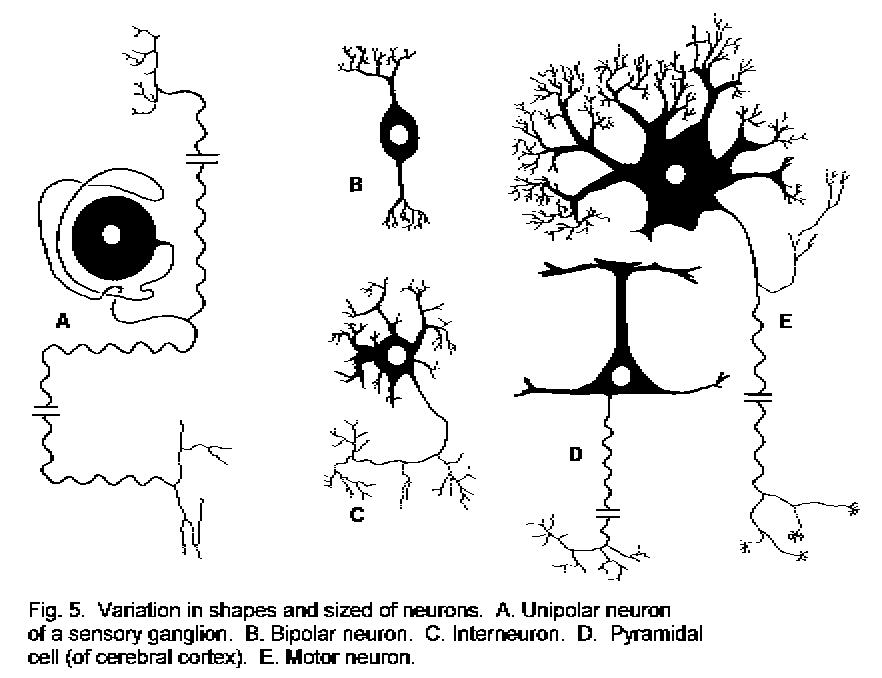
Neurons.
The cells that do the rapid communicating in the nervous system are neurons. These vary greatly in
form
(Picture of some types of neuron), but most types have long cytoplasmic
extensions: dendrites typically conduct towards the cell body, and the single axon typically
carries signals away from the cell body. The primary sensory neuron (dorsal root ganglion cell) differs from
this typical form in being unipolar; the single process is an axon, and it conducts both towards and
away from the cell body.
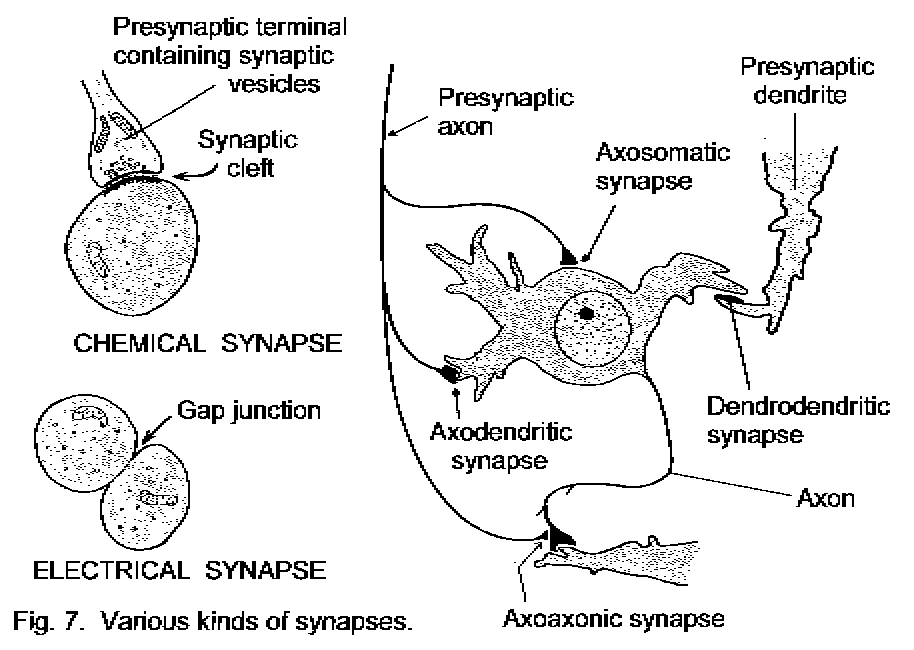
Grey matter, synapses and white matter.
Grey matter contains cell bodies of neurons in the CNS. It is pink because it contains many capillary
blood vessels. It goes grey when the tissue is preserved in formaldehyde. Grey matter also contains dendrites
and axons. Synapses are functional connections between neurons, typically from end branches of axons to
dendrites. Synapses occur in grey matter (and also in autonomic ganglia).
(Diagram of some different sorts of synapse)
Myelin and nerve fibres.
Long axons in the CNS have myelin sheaths, formed from concentric layers of surface membrane derived
from the ensheathing glial cells (oligodendrocytes). Conduction of signals occurs much more rapidly in a
myelinated axon than in one that lacks a myelin sheath. Tissue consisting largely of myelinated axons in the CNS
is called white matter; it is pale in fresh and in fixed tissue. In the PNS myelin sheaths are formed by
the Schwann cells.
Myelin sheath formation in a peripheral nerve
A nerve fibre is an axon together with its myelin sheath.
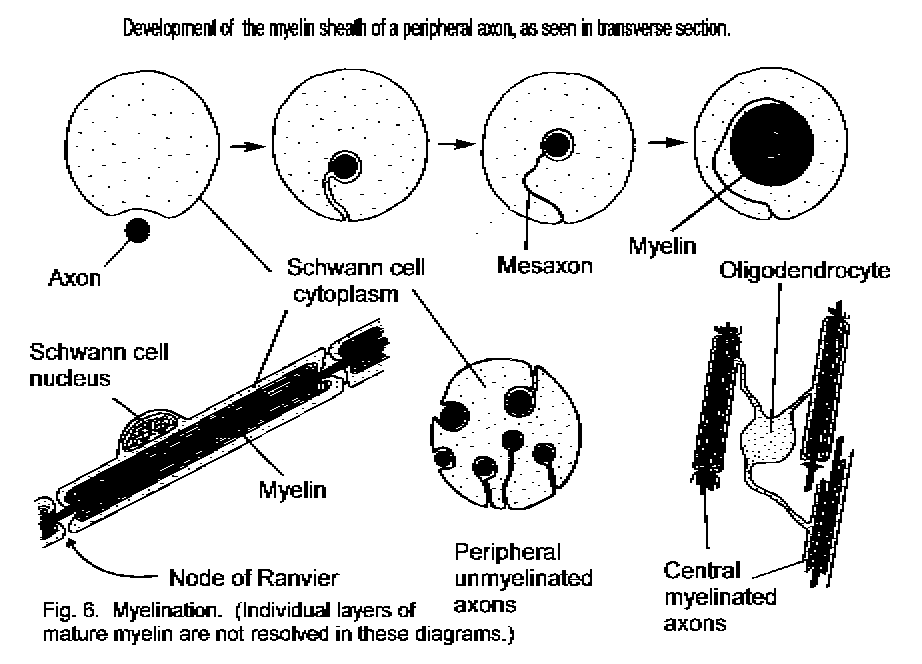
An unmyelinated fibre is an axon that has no myelin sheath and therefore conducts slowly.
Neuroglia.
Other types of cell are also present in nervous tissue, and in the CNS these cells outnumber the neurons by 10:1.
These are the neuroglial cells. In the PNS satellite cells accompany neuronal cell bodies in
ganglia, and Schwann cells ensheath axons and form their myelin sheaths. In the CNS oligodendrocytes form
myelin, and astrocytes fill in all the spaces between neurons and their processes. Microglia are small
cells throughout the nervous system that have functions similar to those of white blood cells of the monocyte/macrophage
series.
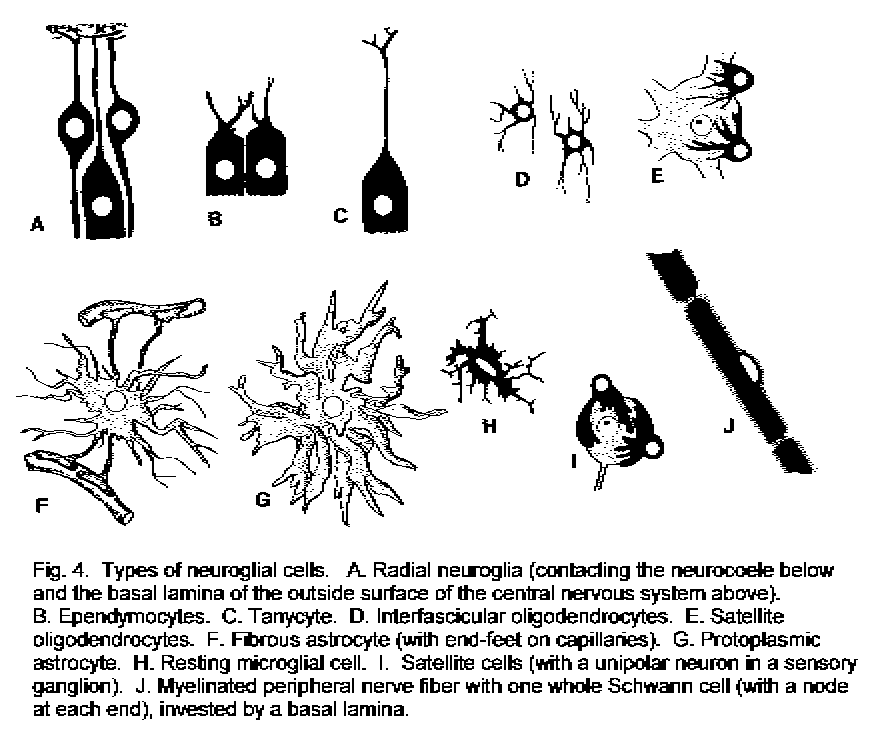
________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
Development of the nervous system and common developmental abnormalities.
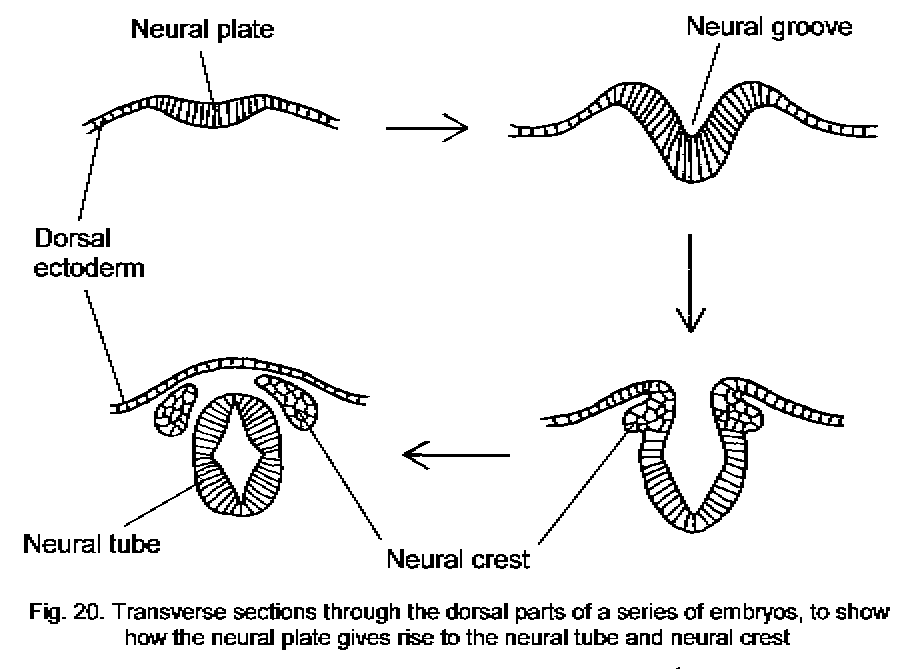
Neurulation
The nervous system develops from the embryonic ectoderm. The neural tube, formed by invagination of the neural plate and fusion of the neural folds, forms the whole CNS (except for the sacral and lower lumbar segments of the spinal cord).
The neural crest, derived from cells of the neural folds, develops into most of
the PNS (neurons and glial cells). In addition, many neural crest cells give rise to
non-neural tissues throughout the body. Some of the neurons in some cranial nerve
ganglia are not of neural crest origin but derive from placodes, which are
localized thickenings of the ectoderm of the embryonic head. The olfactory
epithelium and the sensory receptors of the inner ear also arise from placodes.
Diagram of the events of neurulation
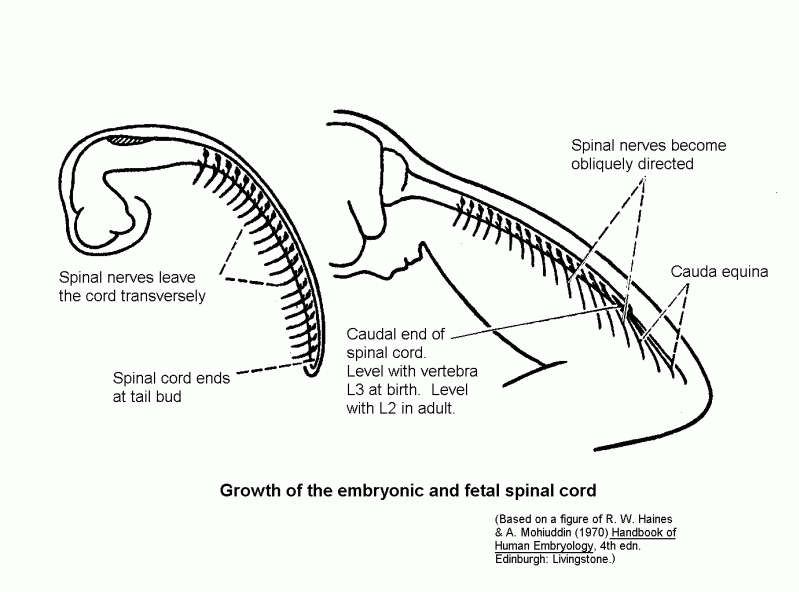
Length of the spinal cord.
The vertebral column elongates more than the spinal cord. Consequently the adult cord
ends at the level of vertebra L2. Below that level the vertebral canal contains the
cauda equina, which consists of the roots of the lower spinal nerves and the
filum terminale. The filum terminale is the stretched-out remnant of the embryonic
tail.
Growth of the spinal cord
Basal and alar laminae.
There are two populations of young neurons in the neural tube, these forming the basal lamina ventrally and the alar lamina dorsally. At the level of the spinal cord, these laminae become, respectively, the ventral and dorsal horns of the spinal grey matter. The ventral horn contains motor neurons whereas the dorsal horn receives the incoming axons of primary sensory neurons.
At the level of the medulla and pons, the roof plate expands to form the
wide, thin roof of the fourth ventricle. Consequently neurons derived from
the alar lamina come to occupy the lateral part of the floor of the 4th
ventricle, and those derived from the basal lamina are medially situated.
(Alar and basal laminae)
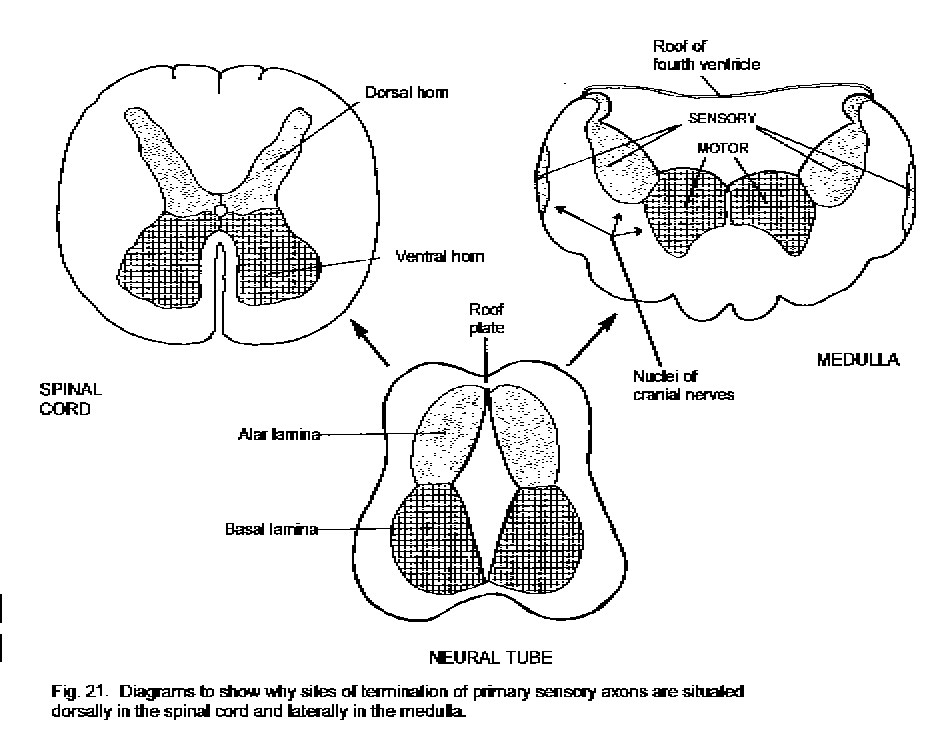
In the medulla and pons, motor nuclei of cranial nerves are therefore located
medially whereas sensory nuclei are located laterally. (A nucleus is a
cluster of similar neuronal cell bodies in the CNS.)
Development of the cerebrum.
Even before the neural folds begin to fuse (Day 18 after fertilization) the future cerebral
hemispheres, brain stem and spinal cord are evident. The rostral and caudal neuropores
close on Days 23 and 27 respectively, and by Day 28 all the major parts of the brain are
in place. (From 8 weeks onward the embryo is called a fetus. The fetal period is characterized
mainly by growth rather than the formation of organs.)
Longitudinal section of 22-day embryo showing the neuropores
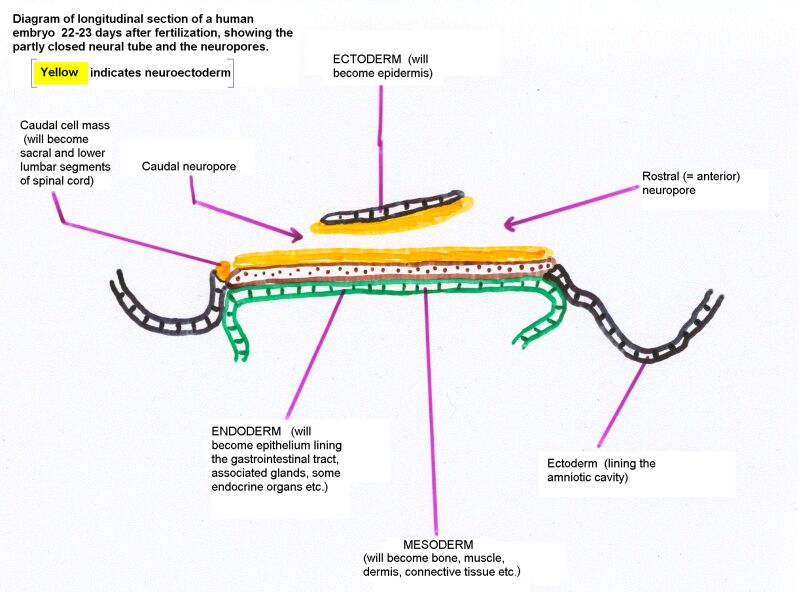
The enlargements of the rostral end (initially the rostral half) of the neural tube are the
hindbrain, midbrain and forebrain. Flexures in the developing brain serve to (a) accommodate
the tubular brain in a round cranial cavity, and (b) allow for the optical axes of the eyes to be
at right-angles to the long axis of the body.
Development of the cerebral hemisphere
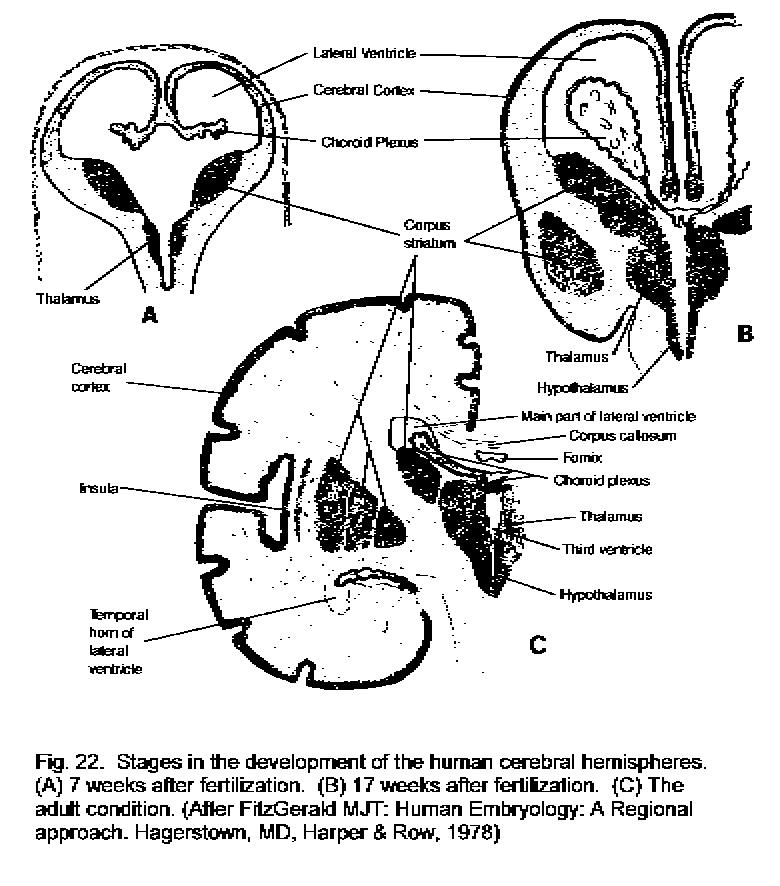
At 11 weeks the main lobes of the cerebral hemisphere are present, but most of the sulci
and gyri have yet to form. Enlargement of the cerebral and cerebral cortex proceeds during
the remainder of the fetal period. Growth of the brain continues into childhood, with most of
the increase in volume being due to myelination of axons and proliferation of dendrites
(associated with increasing numbers of synapses).
Progeny of neural crest cells.
Neural crest cells that remain alongside the vertebral column become the neurons and glial cells of sensory and autonomic ganglia. Those that migrate along the courses of growing axons become the Schwann cells of nerve roots and nerves and the cells associated with specialized peripheral nerve-endings.
Other neural crest cells migrate more extensively. Some become the endocrine cells of the adrenal medulla; others form the melanocytes in the skin. Many cranial neural crest cells differentiate into bones and muscles of the head.
Common abnormalities of development of the CNS.
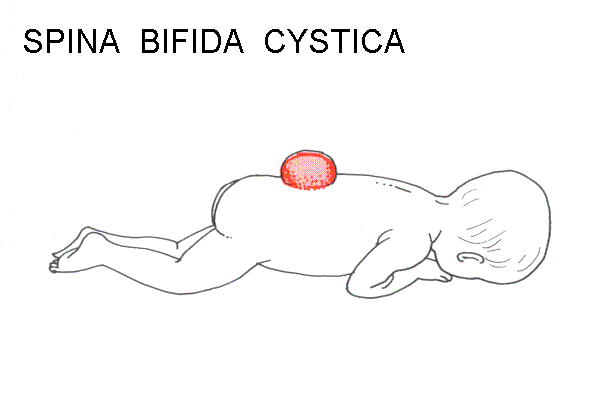
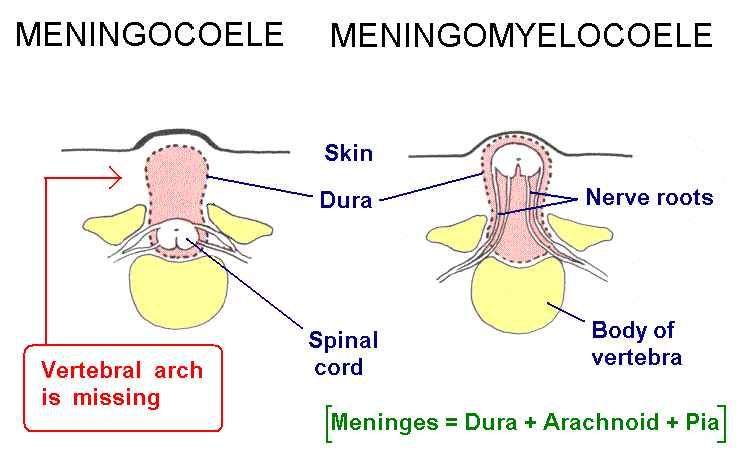
Failure of neuropore closure: Anencephaly if it's the anterior neuropore; myeloschisis if it's the posterior neuropore that fails to close. Most types of spina bifida are due to failure of development of vertebral laminae in the lumbar region. The overlying skin and muscles may be present (spina bifida occulta) or they may be missing (meningocoele or meningomyelocoele). Some drawings of developmental abnormalities follow.
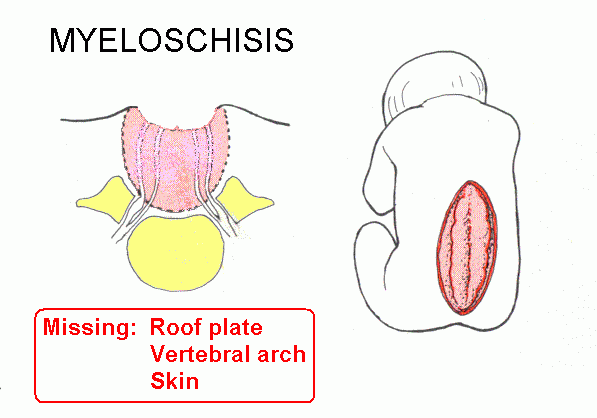
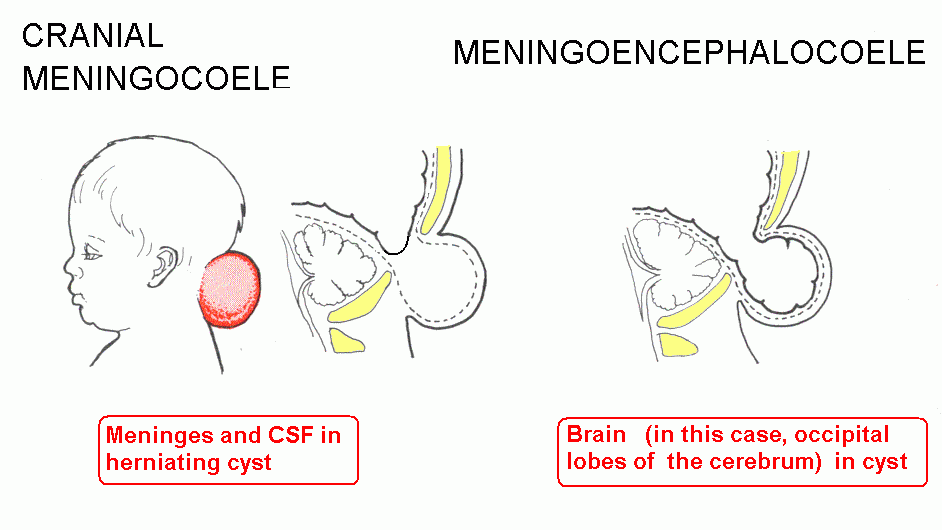
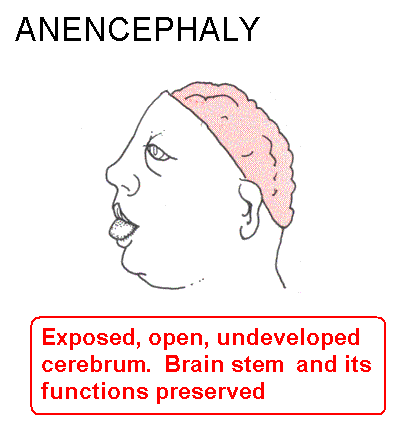
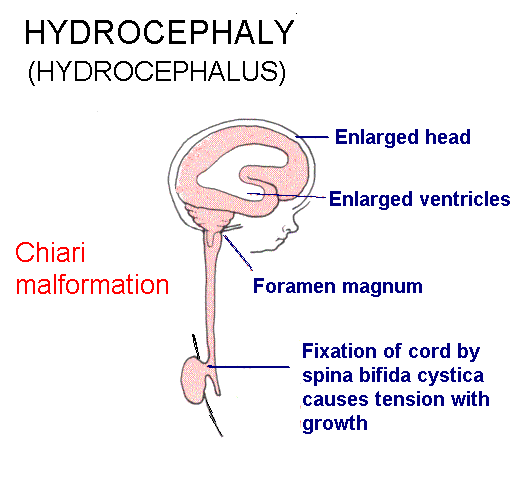
Internal hydrocephalus: Cerebrospinal fluid (CSF) fails to escape from the ventricles, which expand. A common type is due to closure of the cerebral aqueduct. There are malformations that include both hydrocephalus and spina bifida. The Chiari malformations are the most common.
_____________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
Organization of the peripheral nervous system: roots, nerves and ganglia.
In man and in all other vertebrate animals, the nervous system has two divisions: the central nervous system (CNS) is contained in the axial skeleton, and the peripheral nervous system (PNS) is distributed through most of the other parts of the body (Plan of the whole human nervous system).
The central nervous system is connected to other parts of the body by nerves. Neuronal cell-bodies outside the central nervous system occur in ganglia (singular, ganglion). The spinal nerves are segmentally organized. Each has a dorsal and a ventral root, separately connected with the spinal cord. The dorsal root bears a ganglion (called either a spinal ganglion or a dorsal root ganglion); the ventral root does not. The dorsal roots are exclusively sensory in mammals, whereas the ventral roots contain the axons of motor neurons and the axons of neurons that control internal organs, blood vessels, and glands.
Paired cranial nerves connect the brain with other structures. The olfactory nerves, concerned with smell, enter the olfactory bulb, which is at the rostral end of the telencephalon. The optic nerves, like the retinas of the eyes, are made of central nervous tissue. They are therefore not real nerves but outgrowths of the brain. The remaining cranial nerves emerge from the brain stem, which consists of the midbrain pons and medulla.
The nervous system develops from embryonic segments, but in the adult state this is obvious only in the connections of nerve roots with the spinal cord.
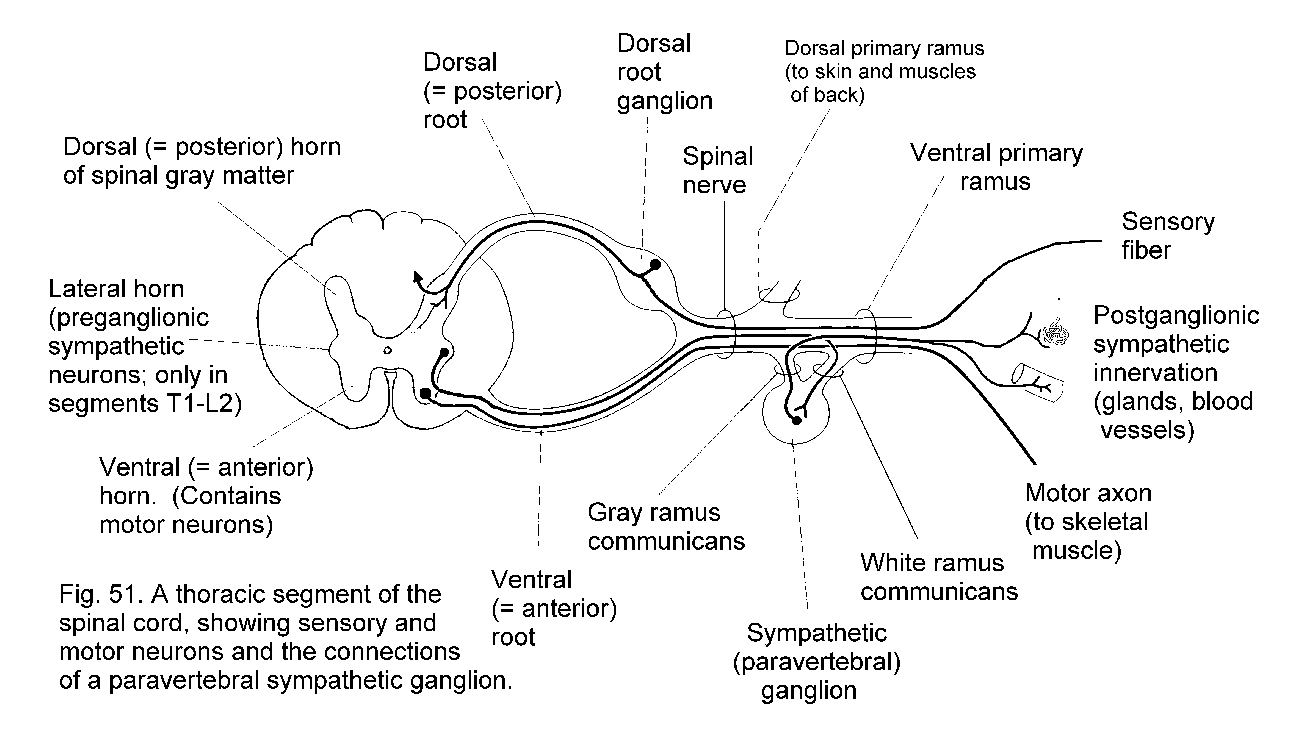
The formation of a spinal nerve is illustrated in Spinal nerve. This diagram also shows structural elements that will be referred to later in this Section.
Spinal nerves have numbers derived
from the vertebrae. The highest spinal nerve penetrates the atlanto-occipital membrane, above
the arch of the atlas, which is the first cervical vertebra or C1. The
second cervical nerve passes between the atlas
(vertebra C1) and the axis (C2). There are 7 cervical vertebrae. The
lowest cervical nerve is therefore C8. Cervical nerves 1 to 7 go
through foramina above
the numbered vertebrae. The roots of nerve C8 pass below
the arch of vertebra C7 and above that of T1. All the thoracic (T1 -
T12), lumbar (L1 - L5) and sacral (S1 - S5) nerves go through foramina below the equivalently numbered vertebrae. To complete the story, a single
coccygeal nerve overlaps with S5 in supplying the perianal skin.
The most obvious consequence of the segmental organization of the spinal nerves is seen in the Dermatomes, which are bands of skin that run horizontally on the trunk and lengthwise on the limbs
(Dermatomes).
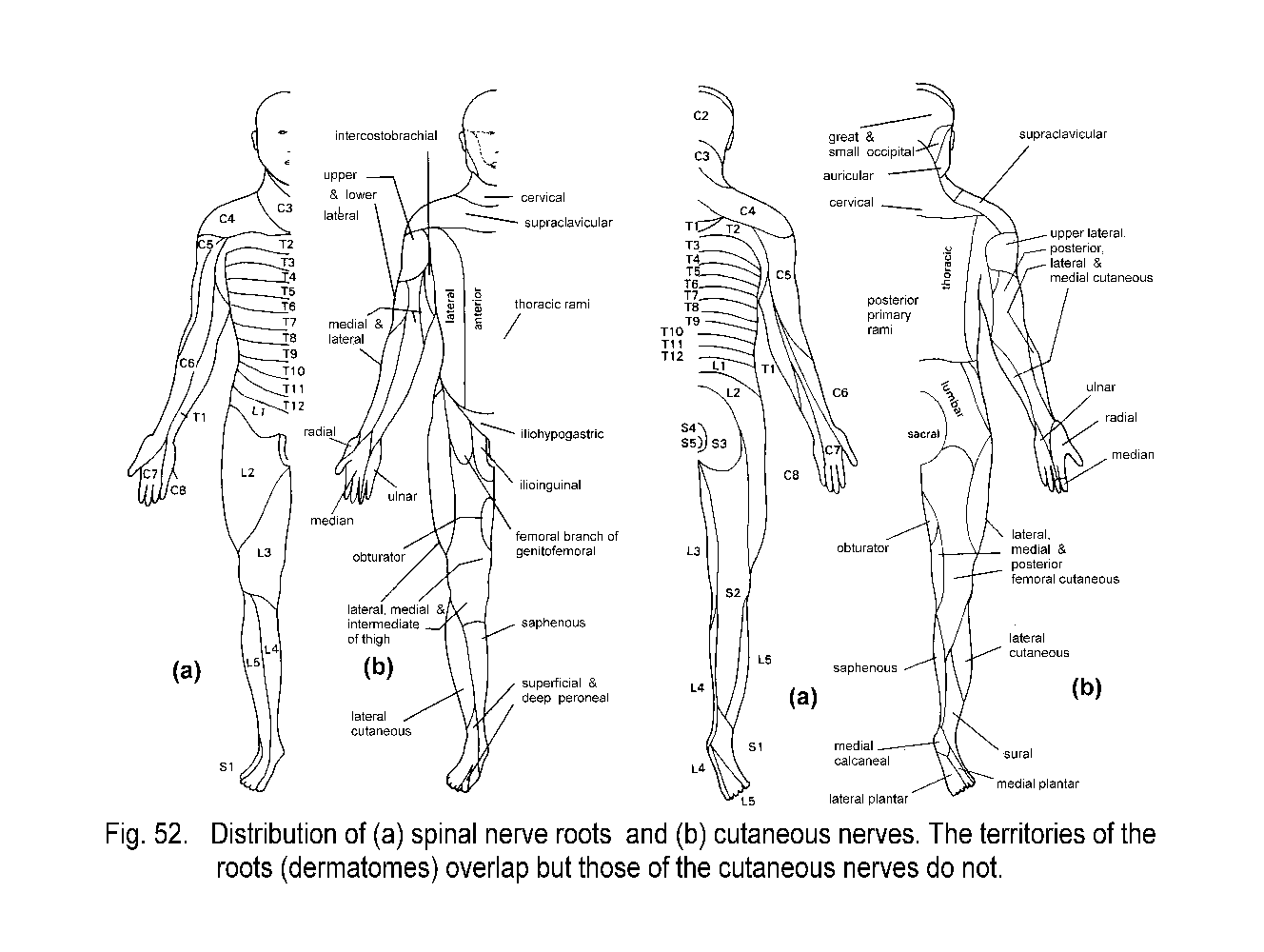
Each dermatome is centered on the distribution of axons from a single
dorsal root ganglion, but each ganglion also supplies skin in the
dermatomes above and below its own level. Consequently, it
is necessary to transect three adjacent dorsal roots or spinal nerves
in order to completely denervate the skin of one dermatome. Transection
of a single spinal nerve, or destruction of its ganglion, diminishes
but does not
abolish sensation in the affected segment of skin. The cutaneous
lesions of herpes zoster, a common virus that infects certain
pain-responsive neurons in individual sensory ganglia, often neatly map
the distributions of
dermatomes and also illustrate the extension of innervation into the
adjacent segments of skin. The nerve supply to the skin of the limbs is
delivered by cutaneous nerves that are formed in limb plexuses
(brachial and lumbosacral) by complex interchanging and mixing of
fibers from different spinal roots. The areas supplied by cutaneous
nerves bear little resemblance to the dermatomes. They are sharply
demarcated, with
little or no territorial overlapping (Dermatomes).
The widely overlapping dermatomes cut across adjacent areas of skin
supplied by cutaneous nerves. A cutaneous nerve lesion, such as an
injury or a mononeuropathy, results in a well defined area of defective
sensation, and anatomical knowledge can be used to identify the
affected nerve.
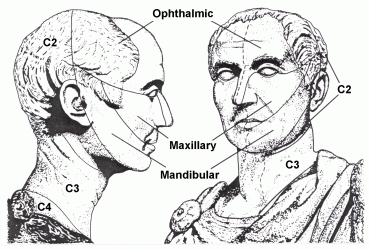
Most of the skin of the head is supplied by the three divisions of cranial nerve V. The areas are sharply demarcated, and therefore do not correspond to dermatomes. Cranial nerves VII, IX and X supply small, overlapping areas of skin of the external ear, and the dermatome of the second cervical nerve includes parts of the head, ear, face and neck. (The first cervical nerve lacks a dorsal root in most people.)
Muscles receive motor and sensory innervation. Most of the muscles of the limbs are supplied nerves formed in the limb plexuses from two or more roots. Table 6 (Segmental landmarks) shows the segmental innervation of a few clinically important muscles. A stretch reflex (tendon jerk) requires the integrity of both the motor and the proprioceptive sensory innervation of the muscle.
| Skin | Muscles (and stretch reflexes) | ||
| C2 | Occipital region of head | ||
| C5 | Tip of shoulder
(The top of the shoulder is in the C4 dermatome) |
||
| C5-C6 | Flexion of elbow (Biceps jerk) | ||
| C6 | Thumb | ||
| C7-C8 | Extension of elbow (Triceps jerk) | ||
| C8 | Little finger | C8-T1 | Small muscles of hand
Oppose thumb to fingers; hold piece of paper between adjacent fingers |
| T4-T5 | Nipple | ||
| T10 | Umbilicus | ||
| L3 | Front of knee | L2-L3 | Quadriceps (Knee jerk) |
| L5 | Big toe | ||
| S1 | Little toe, heel | S1-S2 | Calf muscles (Ankle jerk) |
| S3 and below | Genitalia and anal area | ||
The vertebral column is longer than the spinal cord,
which ends at the level of the upper border of vertebra L3 in the
newborn and at the upper border of vertebra L2 in the adult. The lower
spinal nerves must therefore
course caudally before passing through their corresponding
intervertebral foramina.
Immediately below the caudal end of the spinal cord, the neural canal
contains the roots of nerves L2-L5, S1-S5 and the coccygeal nerve.
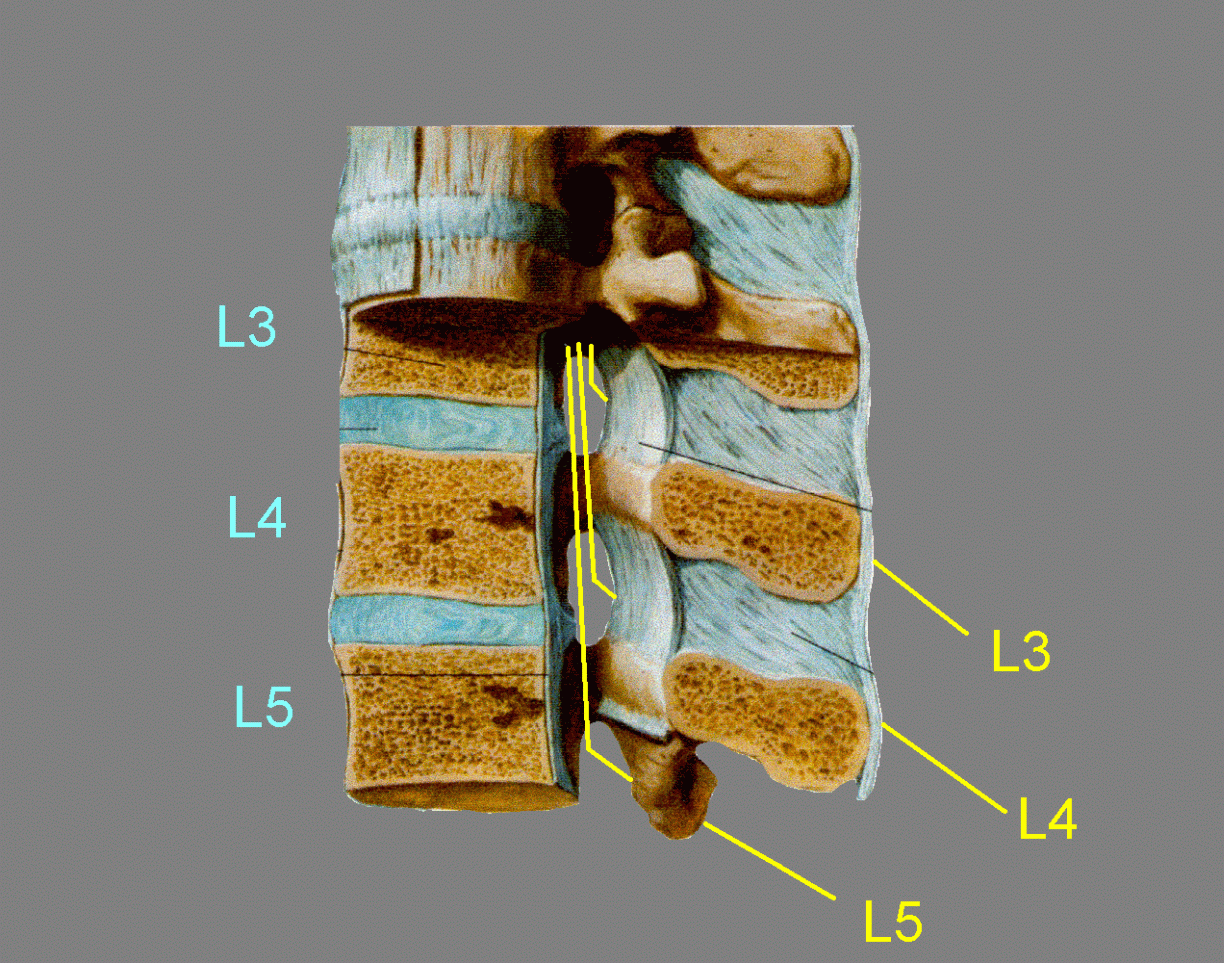
A lesion arising from the axial skeleton, such as a herniated intervertebral disk or tumor tissue from a vertebral body or pedicle, can press on the spinal cord or spinal nerves. The consequences depend on the level of the involved disk or vertebra. In the cervical and upper thoracic spine there is little discrepancy between the spinal segments and the vertebrae. There is little free space in this part of the neural canal, so a lesion is likely to impinge on the cord as well as on a spinal nerve. The body of vertebra T10 is level with spinal cord segment T11. Below this level, the discrepancy between vertebral and spinal levels increases rapidly, because the lower lumbar and the sacral segments of the spinal cord are much shorter than the cervical and thoracic segments. All the spinal cord segments below T11 are in the range of just three vertebrae, T12, L1 and L2.
The foramina are above the levels of the intervertebral disks. Consequently, a herniated disk below C7 cannot compress its own segmental nerve; it presses on the nerve one or two segments lower. For example, an L4-5 disk herniation commonly compresses spinal nerve L5 or S1, causing pain and other sensory abnormalities in the appropriate dermatomes (see Dermatomes figure).
| Functional components | ||||
| Cranial nerve | Motor (= supplying skeletal muscle) | Preganglionic parasympathetic² | General sensory (skin, mucous membranes) | Special senses |
| I Olfactory | Smell | |||
| II Optic | Vision | |||
| III Oculomotor | Eye movements other than those mediated by IV & VI. Elevation of upper eyelid | Constriction of pupil (ciliary ganglion) | ||
| IV Trochlear | Certain downward eye movements | |||
| V Trigeminal | Muscles that open and close the mouth; tensor tympani muscle of middle ear | Skin of face; mouth, teeth, nose, sinuses, dura mater of anterior and middle fossa | ||
| VI Abducens | Abduction of eye | |||
| VII Facial | Muscles of face; stapedius muscle of middle ear | Lacrimal and nasal glands (pterygopalatine ganglion); sublingual & submandibular salivary glands (submandibular ganglion) | Part of external ear and tympanic membrane | Taste: palate & anterior two thirds of tongue |
| VIII Vestibulocochlear: Vestibular Cochlear |
Equilibration Hearing |
|||
| IX Glossopharyngeal | Stylopharyngeus muscle | Parotid gland (otic ganglion) | Pharynx, middle ear, posterior third of tongue | Taste: posterior third of tongue |
| X Vagus | Muscles of larynx & pharynx | Slows heart (cardiac ganglia); increases gastric acid secretion and empties stomach (enteric nervous system) | Larynx, trachea, oesophagus, dura of posterior fossa; part of external ear and tympanic membrane | Taste: epiglottis |
| XI Accessory³ (Spinal component) | Trapezius and sternocleidomastoid muscles | |||
| XII Hypoglossal | Muscles that move the tongue | |||
Footnotes to Table 7
¹ Afferent fibers in IX and X are of great importance for regulation of cardiovascular and respiratory function, but they do not give rise to conscious sensations, and the physiological functions are not usually disturbed by unilateral lesions that affect the nerves or their central connections.
² The names of the parasympathetic ganglia are indicated in parentheses after the functions.
³ The small cranial root of XI carries motor axons destined mostly for the larynx. These cross over into X by way of a communicating branch, as the two nerves pass through the jugular foramen in the base of the skull. The fibers of the spinal root have their cell bodies in segments C1-C5 of the spinal cord.
Skeletal muscles are supplied by motor neurons whose cell bodies are in the spinal cord (anterior horn) or brain stem (motor nuclei of cranial nerves). In contrast, glands, cardiac muscle, and the smooth muscle of blood vessels and internal organs are supplied by neurons in ganglia of the autonomic system. These ganglia receive afferent preganglionic fibers, which are the thinly myelinated axons of neurons in the spinal cord or brain stem. The neurons in the ganglia have unmyelinated axons, the postganglionic fibers that innervate smooth and cardiac muscle and secretory cells. There are three divisions of the autonomic system: sympathetic, parasympathetic and enteric.
The ganglia of the sympathetic system are the chains of paravertebral ganglia that lie on the lateral aspects of the bodies of the vertebrae, and also the preaortic or collateral gangliaassociated
with the branches of the aorta that supply abdominal organs. There is a
sympathetic chain ganglion for every spinal nerve. Postganglionic
fibers enter the nerve by way of a gray ramus communicans (Formation of spinal nerve)
and are distributed to blood vessels, sweat glands and the little
muscles that move hairs. Blood vessels of the skin constrict in
response to their sympathetic
supply, whereas those in muscles dilate. Some of the ganglia for the
nerves C1 to T1 are fused; consequently there are only three cervical
sympathetic ganglia. In most people there is a stellate ganglion, formed from
inferior cervical (C7-C8) and first thoracic ganglia. The middle cervical ganglion is connected with nerves C5 and C6. Postganglionic fibers from the large superior cervical ganglion
(C1-C4) accompany the
carotid artery and its branches. Some enter the eye, where they supply
the dilator pupillae muscle of the iris. Others supply smooth muscle
within the upper eyelid1. All three cervical ganglia send
postganglionic fibers
into cardiac nerves, which run alongside the common carotid
artery and aorta and supply the muscle of the heart. Increased activity
of the sympathetic system increases the rate and force of contraction
of the heart.
Preganglionic sympathetic neurons are present only in spinal cord
segments T1 to L2, where they occupy the lateral horn of the gray
matter. Their myelinated axons constitute the white rami communicantes (see figure),
which are associated only with nerves and ganglia T1-L2. Preganglionic
fibers destined for sympathetic ganglia above and below these levels
pass rostrally and caudally in the
sympathetic trunk, which interconnects all the ganglia of the
sympathetic chain. Some preganglionic fibers pass through thoracic
sympathetic ganglia and emerge as the roots of the greater (T5-T9),
lesser (T10-T11)
and lowest (T12) splanchnic nerves. These nerves pass behind
the diaphragm and end in the preaortic ganglia. Some go to the adrenal
medulla (which is a sympathetic ganglion modified to secrete its
transmitter into
the blood). The efferent axons from the preaortic ganglia accompany
blood vessels to abdominal organs, where most end by synapsing with
neurons of the enteric nervous system (see below).
In the absence of active sympathetic innervation of the eye,
orbit and face, the pupil constricts (unopposed parasympathetic
action), the upper eyelid droops partly, but can be raised voluntarily
(intact oculomotor
nerve innervation) and sweating does not occur on the affected side of
the face. These changes (Horner's syndrome)
can result from damage to the superior cervical ganglion, its pre- or
postganglionic fibers, (which
pass through the stellate ganglion), or to transection of descending
axons in the lower brain stem or spinal cord that control the
preganglionic neurons.
Parasympathetic ganglia are found in the head, connected with certain cranial nerves, and associated with the walls of thoracic and pelvic viscera. Preganglionic fibers leave the brain stem in cranial nerves III, VII, IX and XI and terminate in cranial parasympathetic ganglia. The neurons in these ganglia supply the structures whose functions are stated in Table 7.The cardiac ganglia receive their preganglionic afferents from the vagus nerve; their neurons supply cardiac muscle cells, principally in the atria. The pelvic splanchnic nerves, branches of S2, S3 and S4, carry preganglionic fibers to the parasympathetic ganglia that supply the detrusor muscle of the urinary bladder and the blood vessels of erectile tissue in the genitalia.
The enteric nervous system consists of thousands of tiny, interconnected ganglia in the walls of the alimentary canal, from esophagus to anus, and of some of its associated structures such as the biliary system and pancreas. These ganglia, which supply the smooth muscle and secretory tissues of the gut, contain several types of neurons, with a wide variety of neurotransmitters. The enteric nervous system can do much of its work independently, but it is modulated by preganglionic fibers from the vagus nerve (to the stomach, small intestine and first half of the colon) and from the pelvic splanchnic nerves (distal colon and rectum). Parasympathetic activity stimulates propulsion of the contents of the gut. Of the vagal fibers that enter the abdomen, a majority end in enteric ganglia of the stomach, and the integrity of this preganglionic supply is essential for acid secretion and for opening of the pyloric sphincter.
Most of the postganglionic sympathetic fibers from the preaortic ganglia synapse with neurons in enteric ganglia, but some contact blood vessels and a few supply intestinal smooth muscle. Activity of the sympathetic system causes constriction of visceral blood vessels and retards propulsion of the contents of the alimentary canal.
__________________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
The spinal cord is shorter than the spinal canal in which it is suspended. Except in the neck, spinal cord segments are rostral to the corresponding vertebrae. The end of the adult spinal cord (the conus medullaris) is level with the 2nd lumbar vertebra.
Lumbar puncture: Cerebrospinal fluid can be sampled by a needle put into the subarachnoid space below the level of the conus medullaris.
The cross sectional area of the central gray matter indicates the number of neurons: largest for segments supplying limbs.
(Different levels)

The cross sectional area of the white matter decreases caudally because there are fewer descending and ascending fibers.
Motor neurons are in the ventral horn; sensory axons enter the dorsal horn and the dorsal funiculi. Preganglionic autonomic neurons are laterally placed, in segments T1-L2 and S2-S4.
Section of a thoracic segment of the spinal cord showing major groups of neurons in the grey matter, and positions of tracts in white matter.
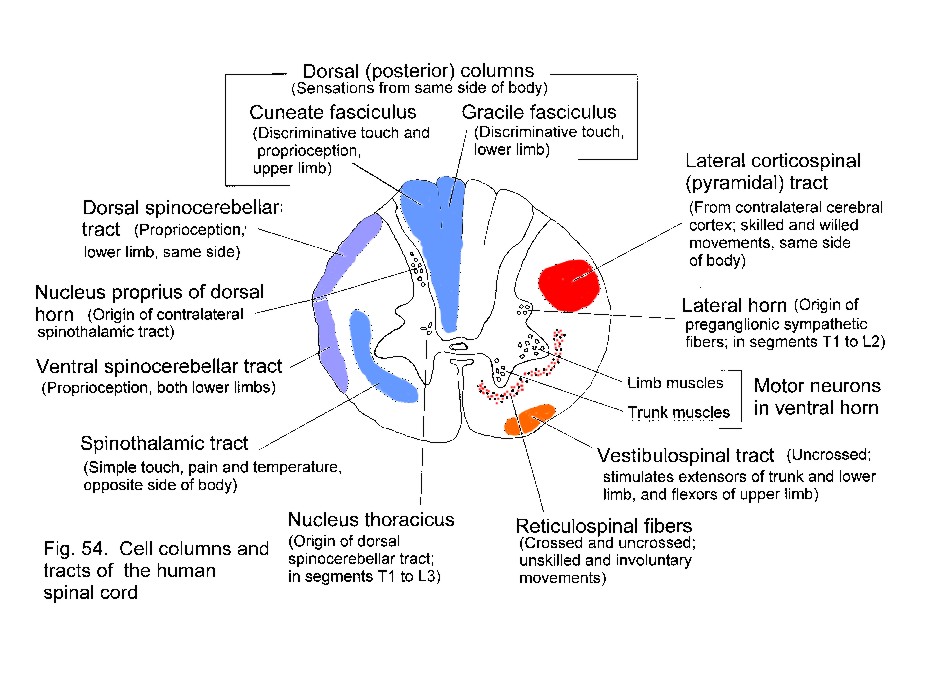
Ascending tracts include the uncrossed gracile and cuneate
fasciculi (from sensory ganglia) and the crossed spinothalamic tract
(from the dorsal horn). These are concerned with different types of
sensation.
Descending motor tracts include the uncrossed vestibulospinal and the crossed lateral corticospinal tract. Hypothalamospinal and some reticulospinal fibers influence autonomic functions.
Lesions
in different parts of the spinal cord produce sensory and motor
abnormalities appropriate to the functions of the tracts that have been
transected. The segmental level of a lesion is indicated by the
affected dermatomes and movements.
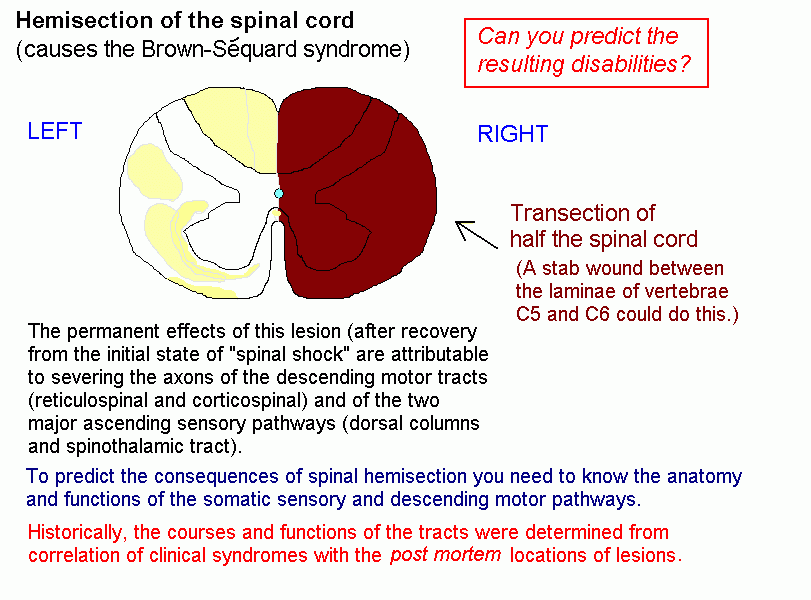
Lesion causing the Brown-Sequard syndrome
Can you determine what the motor and sensory deficits are, for parts of the body below the segmental level of the lesion?
Spinal reflexes.
A reflex has afferent and efferent limbs. The simplest one is the stretch reflex,
see diagram. This includes only one synaptic
interruption. Branches from the axon of the proprioceptive neuron
directly contact motor neurons for the same muscle.
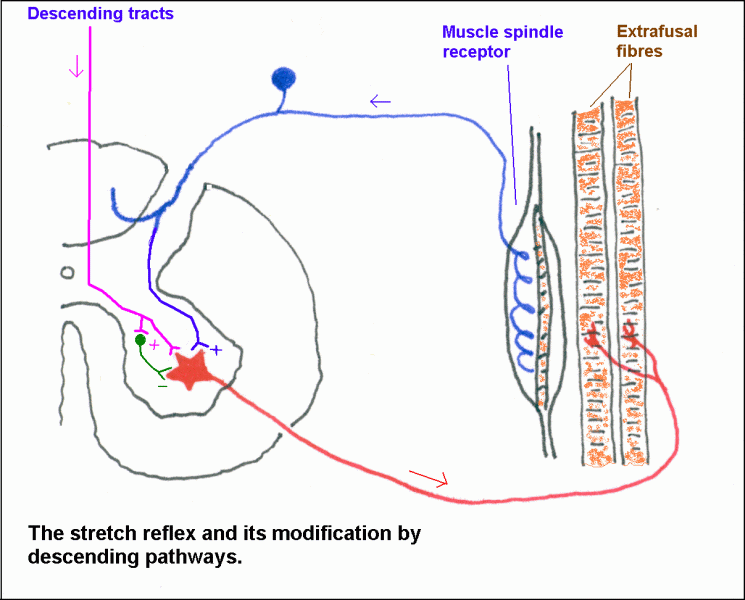 Descending
pathways generally suppress stretch reflexes. If descending motor
pathways are severed, by disease or injury, the resulting spasticity of the paralyzed muscles is due to uninhibited stretch reflexes.
Descending
pathways generally suppress stretch reflexes. If descending motor
pathways are severed, by disease or injury, the resulting spasticity of the paralyzed muscles is due to uninhibited stretch reflexes.
__________________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
The cerebral cortex: anatomical landmarks and location of primary sensory and motor areas.
It is easy to get bogged down in the names and positions of sulci and gyri. Start with those that are easily seen, and don't bother with ones that are not listed here.
| Longitudinal (interhemispheric) fissure | Between the cerebral hemispheres. Its floor is the corpus callosum. | |
| Lateral sulcus (sylvian fissure) | Separates temporal lobe from frontal and parietal lobes | |
| Insula | Landmark for underlying lentiform nucleus | |
| Central sulcus | Landmark for primary motor and somatic sensory areas | |
| Frontal lobe | ||
| Precentral gyrus | Primary motor area | |
| Inferior frontal gyrus | Includes Broca's expressive speech area | |
| Prefrontal cortex | Complex functions involving foresight | |
| Anterior cingulate cortex | Memory; movement | |
| Premotor area | Control of movement | |
| Supplementary motor area | Initiation of movements | |
| Frontal eye field | Saccadic eye movements (not pursuit or vergence) | |
| Parietal lobe | ||
| Postcentral gyrus | Primary somatosensory area | |
| Superior parietal cortex | Somatosensory association area; lesions cause apraxia, neglect | |
| Inferior parietal cortex | Higher order association areas for language, calculation; lesions cause receptive aphasia | |
| Posterior parietal cortex | Higher order association area for vision; eye-field for pursuit movements | |
| Occipital lobe | ||
| Calcarine sulcus
(and the adjacent gyri) |
Primary visual area | |
| Remainder of the occipital lobe | Visual association cortex | |
| Temporal lobe | ||
| Inferior and inferolateral
regions |
Highest order visual association area, including memories of complex scenes | |
| Superior temporal gyrus: | ||
| Anterior & middle part
of superior surface |
Primary auditory area | |
| Middle and posterior parts | Auditory association area; also called Wernicke's area; part of the association area for language | |
| Parahippocampal gyrus: | ||
| Uncus | Primary olfactory cortex | |
| Entorhinal area | Includes primary and association cortex for olfaction. Afferents from all sensory association areas. Efferents to the hippocampus. | |
| Fusiform gyrus (lateral
to collateral sulcus) |
Includes visual association cortex for remembering people's faces | |
| Hippocampal formation
(subiculum, hippocampus, dentate gyrus etc) |
Integration of sensory experiences and formation of memories | |
| Medial surface of hemisphere | ||
| The cingulate gyrus,
cingulate sulcus and parieto-occipital sulcus are conspicuous landmarks. |
The corpus callosum interconnects symmetrical areas of cerebral cortex of the two cerebral hemispheres | |
Functional areas of the cerebral cortex
Here is the Map
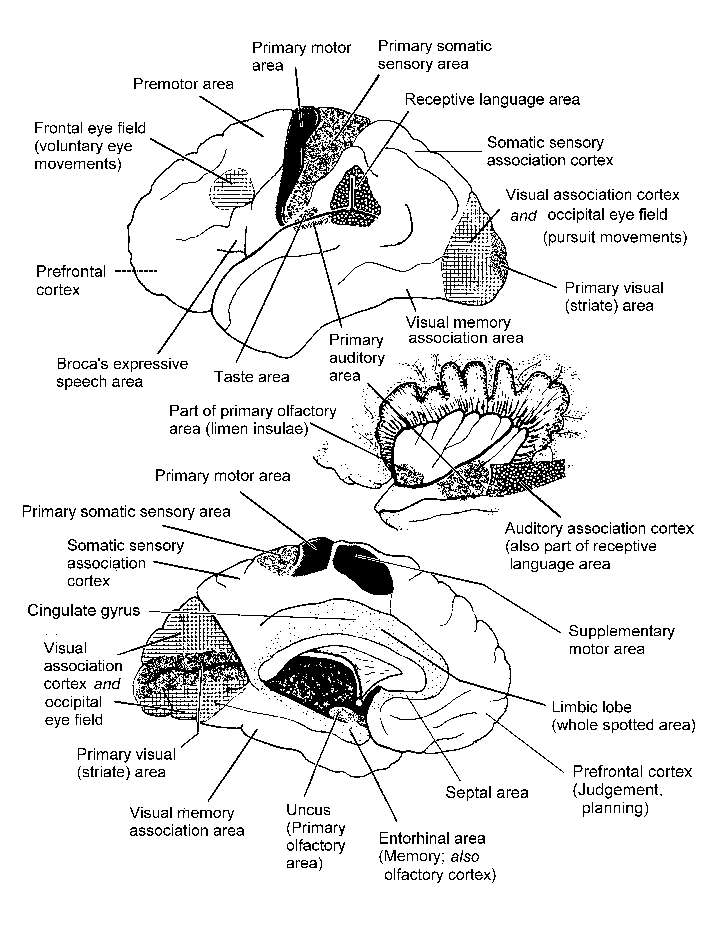
.
.
.
.
.
.
.
.
.
.
.
.
This drawing shows the Somatosensory and motor homunculi
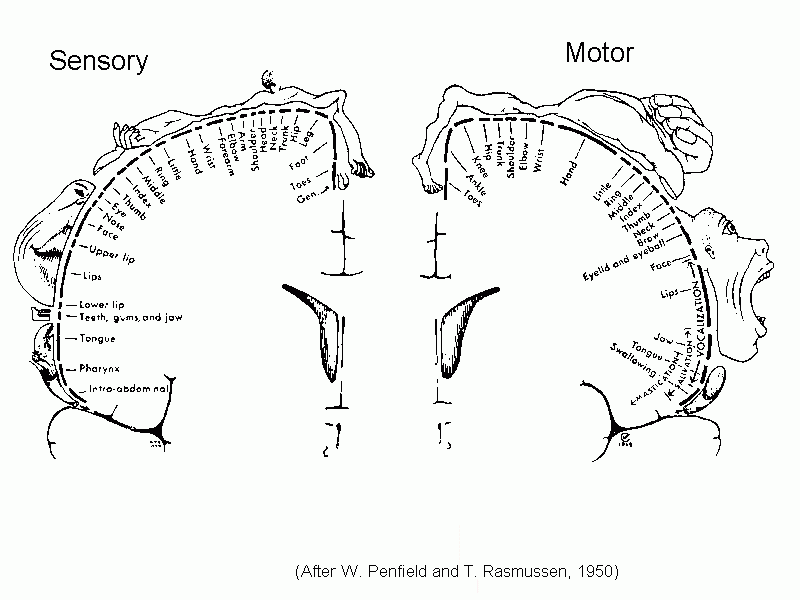
This classical picture of the homunculi, based on the work of Wilder Penfield
in the 1930s and 1940s has recently been found to contain an error. The face should be upside down, but it is
shown the right way up!
[See Servos P, Engel SA, Gati J & Menon R (1999). fMRI evidence
for an inverted face representation in human somatosensory cortex. NeuroReport
10(7):1393-1395.]
__________________________________
BRAIN STEM ANATOMY
(Click here)
__________________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
INTERNAL ANATOMY OF BRAIN STEM AND DIENCEPHALON
This subject is covered in the practical classes. Students need to know the anatomy as shown
by dissection and should also be able to identify the structures seen in:
(a) A horizontal slice at the level of the insula and internal capsule.
(b) A coronal slice at the level of the insula and internal capsule.
__________________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
Arteries of the central nervous system and the regions they supply.
The vasculature of the CNS is adequately described in the "Manter & Gatz" textbook, with rather more details than are needed for Anatomy 530a.
The arteries at the base of the brain anastomose in the circle of Willis (circulus arteriosus, arterial circle).
The arteries forming the circle, and
the branches that supply the brain, can be summarized in a simple
thumbnail diagram, which is easily learned.
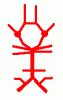
Click here for a labelled version of this simple diagram.

Click here for an unlabelled colour photo that shows most of the arteries at the base of the brain.
The brain is supplied by 4 arteries. On each side:
The internal carotid artery enters the skull through the carotid canal
and then traverses the cavernous sinus (= lateral parasellar compartment).
The vertebral artery (a branch of the thyrocervical trunk of the subclavian artery)
passes through foramina in the transverse processes of cervical vertebrae, pierces the
atlanto-occipital membrane, and enters the cranial cavity by way of the foramen magnum.
Picture of the inferior surface of the brain with arteries labelled.
In this illustration the groups of central arteries that supply the internal parts of the cerebral hemispheres are clearly
shown but are not labelled.
The central arteries are proximal branches of the three cerebral arteries and branches of the posterior communicating arteries.
The lenticulostriate arteries (from the middle cerebral) supply the internal capsule.
The three cerebral arteries supply the cortex:
Anterior cerebral: Medial surface and top of hemisphere, excluding occipital lobe.
Posterior cerebral: Occipital lobe, inferior surface of temporal lobe; also amygdala, hippocampal formation.
Middle cerebral: Most of the lateral aspect of the hemisphere - frontal, parietal and temporal lobes.
Also, in the subcortical white matter, association fibres (notably the arcuate fasciculus) and
projection fibres (notably the geniculocalcarine tract).
The vertebral and basilar arteries send branches to the brain stem and cerebellum. The posterior inferior cerebellar artery supplies the lateral part of the mid- and upper medulla in addition to the cerebellum.
The caudal medulla and spinal cord are supplied by branches from the vertebral arteries.
The labyrinthine artery, usually a branch of the anterior inferior cerebellar, supplies the inner ear.
Note
the position of the oculomotor nerve, medial to the uncus and also in
the angle between the posterior cerebral artery (which supplies
structures above the tentorium cerebelli) and the superior cerebellar artery (which supplies structures below the tentorium).
If the cerebrum is pushed downward (by an expanding intracranial
haemorrhage or tumour, for example), there is pressure on the nerve (What happens?), and the posterior cerebral arteries can be stretched over the free edge of the tentorium and occluded. What are the expected effects of bilateral PCA occlusion?
Venous drainage and the absorption of CSF.
Click here for a full sized image
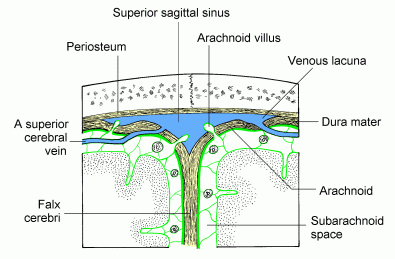
Superficial cerebral veins traverse
the subarachnoid space and then pierce the dura as they enter
intracranial (dural) venous sinuses. The diagram shows 2 veins entering
the superior sagittal sinus.
Head injury (rapid rotation of brain within cranium) can tear veins as the traverse the dura, causing subdural haemorrhage.
The arachnoid granulations allow cerebrospinal
fluid to flow into the venous blood of the sinus but prevent back-flow
of blood into the subarachnoid space (a valve mechanism).
__________________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
Neuronal signals from skin and deeper structures are segregated in the spinal cord. Transmission to the thalamus and cerebral cortex may occur through the spinothalamic tract or through the dorsal funiculi and medial lemniscus.
For pain, temperature and the less discriminative aspects of touch, neurons in the dorsal horn have axons that cross in the spinal cord and ascend as the spinothalamic tract, which is laterally situated in the spinal cord and brain stem.
For discriminative touch and for conscious proprioception, the axons of
primary sensory neurons ascend ipsilaterally in the dorsal funiculus
(either gracile or cuneate fasciculus) and end in the gracile or cuneate
nucleus. Fibers arising in these nuclei cross in the medulla and ascend
in the medial lemniscus, which is near the midline in the medulla and
shifts to a lateral location in the midbrain.
For conscious proprioception from the lower limb there is an additional pathway,
involving the dorsal spinocerebellar tract and nucleus Z in the medulla.
|
|
Click here to download sompaths.exe (311073 bytes). This is a free-standing executable slide-show - the one used for the lecture. (This needs MS Windows 95 or later. Sorry about that. If you use an older computer, I can provide plenty of DOS-based software that's much better than this Windows stuff, but some of it won't run on computers made since 1998-99. Ask me if you're interested. None of my downloadable programs are for Mac computers.) |
The differences between the two main ascending somatosensory pathways are important functionally and clinically.
|
The medial lemniscus system (= dorsal column system = dorsomedial system) is for discriminative touch and conscious proprioception.
The pathway ascends ipsilaterally in the dorsal white matter of the cord, and crosses in the caudal (closed) medulla. In the medulla the medial lemniscus is close to the midline; the tract moves laterally as it ascends through the pons and midbrain. |
The spinothalamic system (= anterolateral system) is for simple touch, warmth, cold and pain.
The pathway crosses the midline immediately and ascends in the lateral and ventral white matter of the spinal cord. The spinothalamic tract maintains a lateral position as it traverses the medulla, pontine tegmentum and midbrain. |
| Both the spinothalamic tract and the medial lemniscus terminate in the ventral posterior nucleus of the thalamus. This thalamic nucleus projects to the primary somesthetic cortex of the postcentral gyrus, where the contralateral half of the body is represented as an upside-down homunculus. |
|
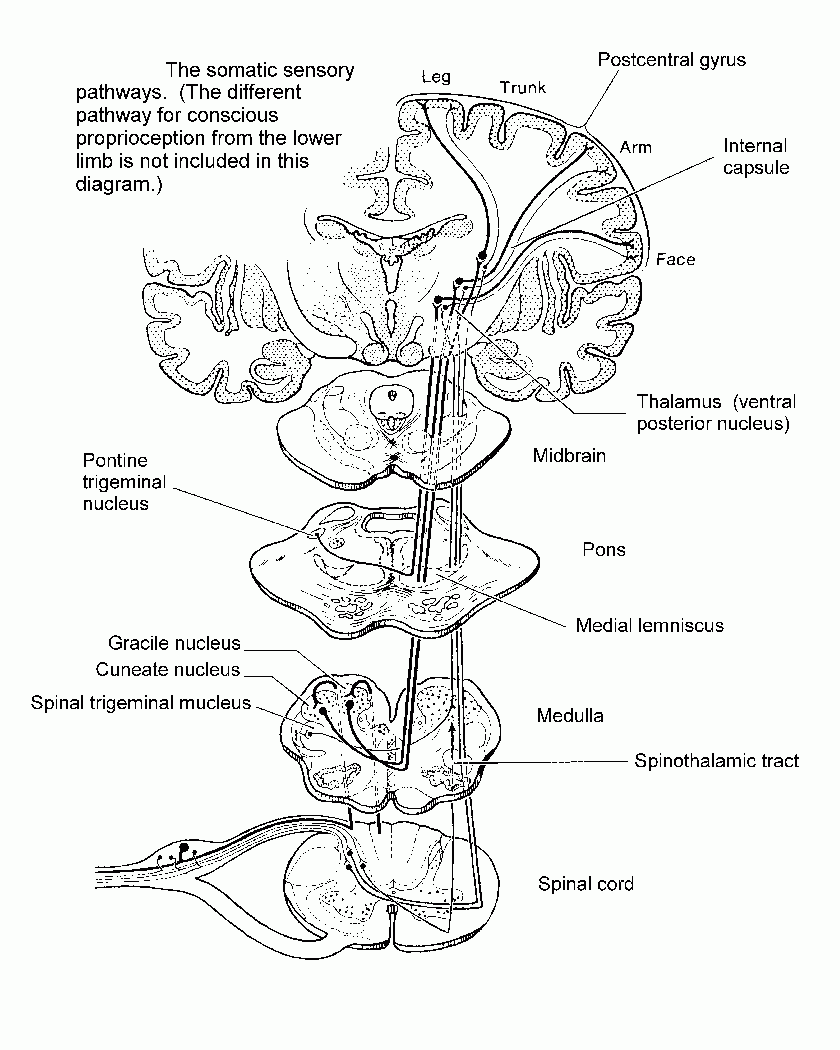
|
Diagram summarizing the somatic sensory pathways |
The somatic sensory pathways for the head involve the trigeminal sensory nuclei and
their projections to the contralateral VPM thalamic nucleus.
Primary afferent fibers for touch end in the pontine trigeminal nucleus.
Pain and temperature fibers descend in the spinal trigeminal tract before ending in
the caudal part of its nucleus. This pathway will be discussed later, in connection with
the central connections of cranial nerve V.
Lesions in the spinal cord and brain stem can affect the somesthetic pathways
separately, causing dissociated sensory loss.
What sensory loss results from a destructive in the
lateral part of the medulla, involving the left spinothalamic tract and sensory
nuclei of the left trigeminal nerve (but sparing other somatic sensory pathways)?
Click here for the answer (but think first!)
The main pathways are supplemented by others, especially for pain, which involve the reticular formation and thalamic nuclei other than the VPL or VPM .
The cerebral cortex is necessary for localizing the source of a painful stimulus and for the recognition of objects by touch.
A note about nuclei in sensory pathways.
The synapses between the main neurons in a pathway are not simply "relays," which would serve
no useful purpose. Through connections from other parts of the CNS, and with the involvement
of local interneurons, the incoming signals are modified for onward transmission.
For example:
In the gracile and cuneate nuclei,
lateral inhibition
(feed-forward and feedback types) sharpens the perception of the most strongly
stimulated part of a receptive field. There is also remote inhibition
by corticobulbar fibres in these nuclei.
In the dorsal horn of the spinal cord, large and small diameter axons contact
inhibitory interneurons and tract cells that send pain signals to the
thalamus. Balance between the two types of input constitutes the
gate control mechanism
for pain.
Descending pathways that influence sensation
The proper interpretation of the outside world and the state of the body itself would be impossible if every impulse in every sensory axon were to be brought eventually to the cerebral cortex. An editing system is necessary, so that the cortex can select the sensory information worthy of conscious attention while leaving more humble duties to the spinal cord, brain stem, and cerebellum. The editing function is carried out by descending fibres that terminate in the sites of origin of the ascending tracts. Some of these descending pathways are shown in a diagram (Click here).
Descending tracts modify activity in the ascending systems at three levels:
1. In the dorsal horn of the spinal gray matter. This is the site of termination of large
numbers of corticospinal fibers, mostly from the postcentral gyrus. Other axons
ending in the dorsal horn come from the reticular formation and from the gracile
and cuneate nuclei. One of the reticulospinal projections, the raphespinal tract,
is notable for inhibiting the upward transmission of signals concerned with
pain. It originates in the raphe nuclei, in the midline of the medulla, and the unmyelinated serotonergic
axons of the tract are lateral to the tip of the dorsal horn.
The raphe nuclei are themselves stimulated by neurons in the periaqueductal
gray matter of the midbrain.
Electrical stimulation of the periaqueductal gray
causes prompt relief of pain and has occasionally been done clinically for this
purpose. A curious observation is that stimulation for a few minutes can produce
analgesia lasting several hours.
2. In the brain stem. Large numbers of fibers descend from the somatosensory
area of the cerebral cortex to the gracile and cuneate nuclei. They are presumed to influence the
medial lemniscus system. Corticobulbar fibers also end in the trigeminal sensory nuclei.
The suffix "-bulbar" refers to the termination
of axons in the brain stem. The term "corticonuclear"
is sometimes used for axons of cortical neurons that end in
nuclei of cranial nerves. However, many descending fibers
that influence cranial nerves end near, but not within, the nuclei.
The less precise term "corticobulbar" is therefore preferred.
3. In the thalamus. The VPL and VPM nuclei project to the first somatosensory area of
the cerebral cortex. These thalamic nuclei also receive input from the same cortical
areas.
(This isn't the only thalamocortical
projection that is reciprocated; they all are.)
________________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
DESCENDING MOTOR PATHWAYS (Click here) for some notes.
________________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
THE CEREBELLUM (Click here) for some notes.
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
THE BASAL GANGLIA (Click here) for some notes.
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
Some cranial nerves concerned with special senses, and their central connections:
I Olfactory system.
The
olfactory system is presented as a 15-minute videotape entitled "The
Rhinencephalon" by D. G. Montemurro. This tape, shown in the lecture,
is also available in the Taylor (Sciences) Library.
The following three images summarize the system.
For Anatomy 530a you do not need to know about cells in the olfactory
epithelium other than the receptor neurons.
_______________________________________
Location of olfactory epithelium, nerves and bulb
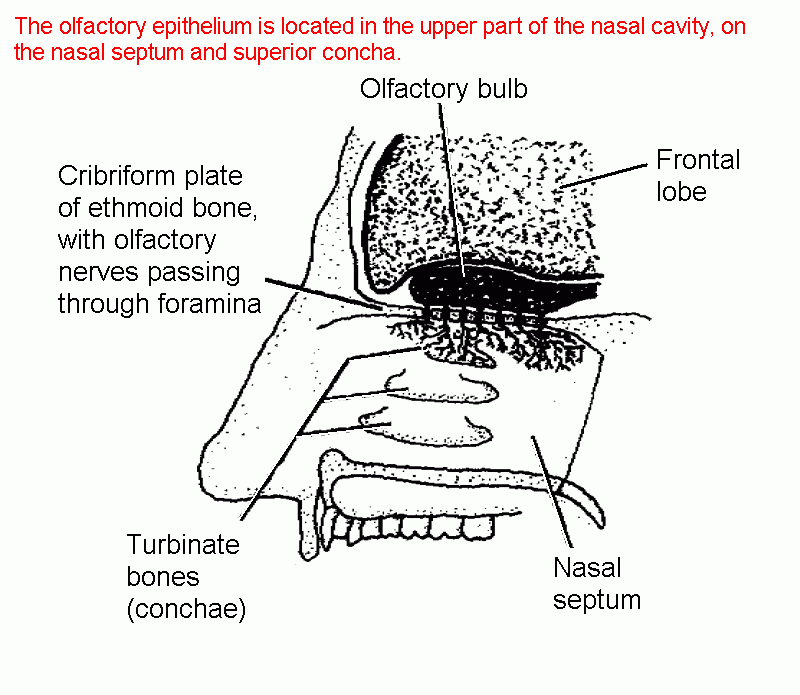
Structure of the olfactory epithelium (annotated diagram)
Destinations of fibres of the olfactory tract
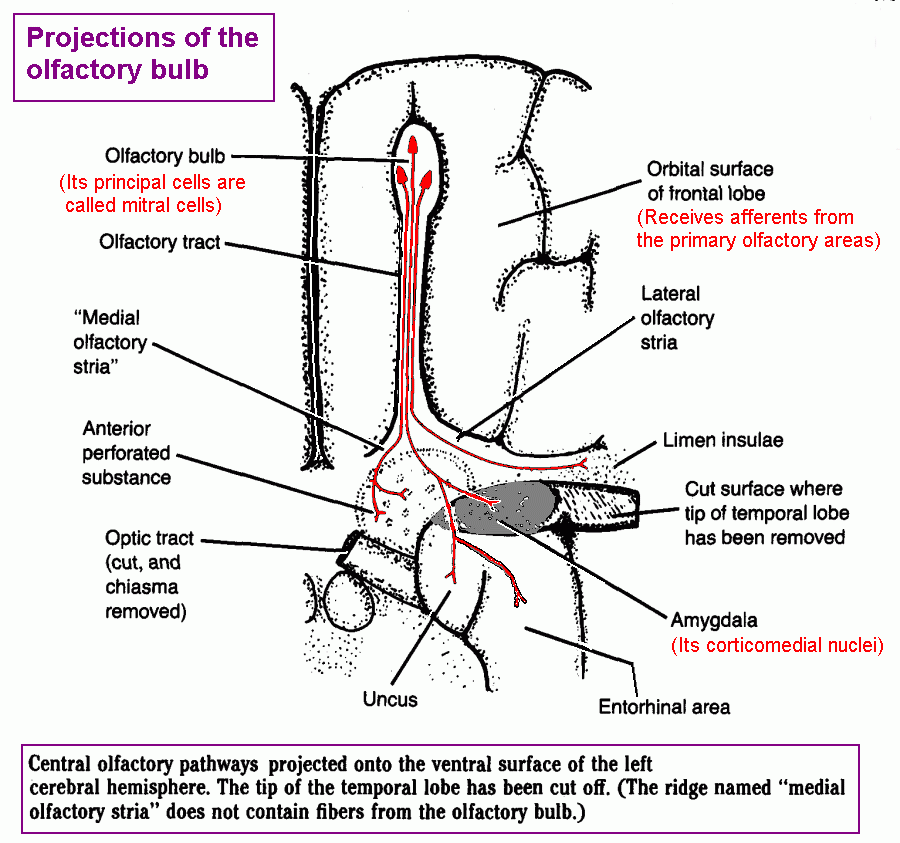
II Visual system.
Damage
to the visual pathways can result in visual field defects, with the
site of a lesion determining which part of the visual fields is blind.
To understand the visual pathways it is first necessary to be familiar
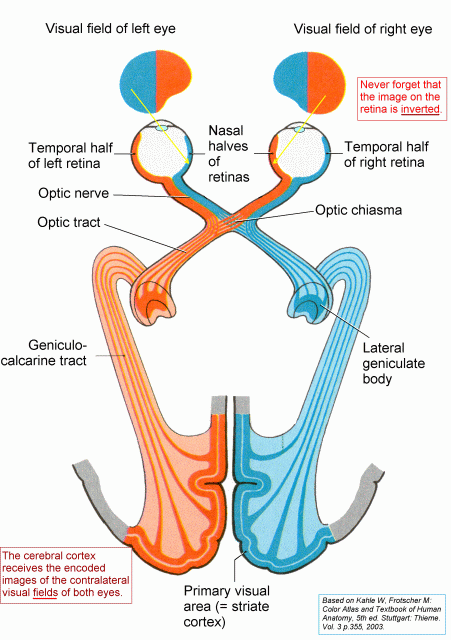
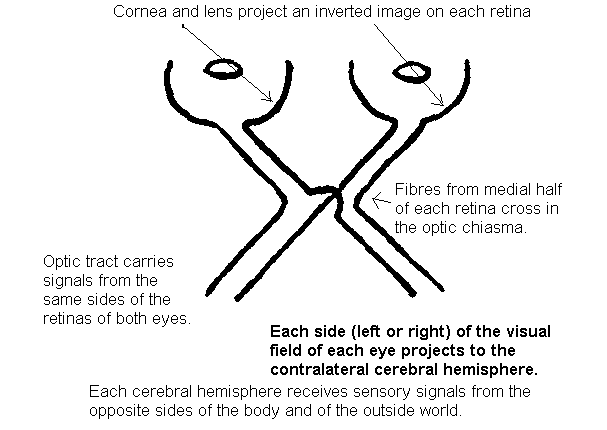
with the geometrical optics of the eye, and to appreciate that (a) images projected onto the retina are inverted, both left-right and top-bottom, and (b)
in man, both eyes are directed forwards, so that almost all the visual
field is projected onto both retinas. It is also necessary to know that
there is retinotopic organization throughout the pathways.
Remember that the visual field is seen by both eyes (because they face forwards) and that an inverted
image of the visual field is projected onto both retinas. It is necessary to know this in order to understand the topographical
representation of different parts of the visual fields in the brain.
In addition to the lateral geniculate body there are at least 4 other destinations
of fibres of the optic tract. These include the pretectal area for the pupillary light
reflex and the superior colliculus which is involved in the control of eye movements.
The lens of the eye is shown in the thumbnail sketch to remind you that images on the retinas are inverted.
The notes below relate to images. Click the links on the right side of the table to look at the images (which are the ones shown in the lecture).
* Meyer's loop contains
geniculocalcarine fibres that are deflected into the temporal lobe by
growth of the temporal horn of the lateral ventricle. These fibres
carry signals from the inferior halves of the contralateral
hemiretinas. What visual field defect results from a destructive lesion
in the right temporal lobe? Try to work this on out yourself, as
did Adolph Meyer (1866-1950), the Austrian ophthalmologist who deduced
the topography of the geniculocalcarine projection.
Anatomy and optics of the eye.
A horizontal section through the left eye, viewed from above, showing the focusing of a real, inverted image on the retina.
Behind the lens, the eyeball is filled by the vitreous body, which is transparent and gelatinous. The space between the lens and the cornea is occupied by aqueous humour,
a liquid secreted by the ciliary processes (anterior to ciliary muscle)
and absorbed into veins at the angle between the iris and the cornea.
The posterior chamber of the eye is between the lens and the iris; the anterior chamber is between the iris and the cornea.
The cells of the choroid contain a pigment, melanin, which prevents
reflections of light within the eyeball. Melanin occurs also in the
iris.
Anatomy and optical properties of the eye

The retina is part of the CNS (outgrowth of the diencephalon), as is the optic "nerve."
The cellular organization of the retina is complicated.
Diagram of retinal structure
[This diagram shows more detail than is required for Anatomy 530a]
The ganglion cells of the retina have axons that pass through the optic nerve to the midbrain and thalamus.
The fibres from the medial (nasal) halves of the two retinas
cross the midline in the optic chiasma and go into the contralateral
optic tracts.
The fibres from the temporal halves of the retinas go into the ipsilateral optic tracts.
The visual pathway (for conscious visual sensation)
This thumbnail drawing is helpful for working out visual field defects.
It can be used to solve all the classical visual field problems other than a Meyer's loop* lesion.
 Thumbnail diagram showing what crosses and what doesn't in the optic chiasma
Thumbnail diagram showing what crosses and what doesn't in the optic chiasma
_______________________________________
VIII (cochlear). Auditory system.
Sounds are detected by a receptor of great structural complexity, the organ of Corti in the cochlea of the inner ear.
Drawing that shows some anatomical features of the ear
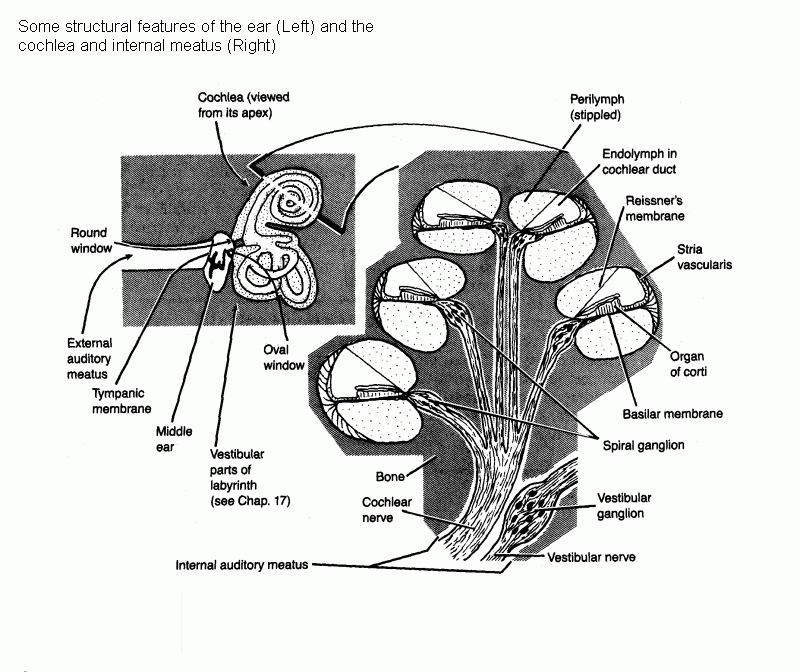
The neurons in the spiral ganglion are bipolar (perhaps best considered to have two axons). Their distal processes receive synaptic input
from cells of the organ of Corti. Their proximal processes form the cochlear nerve (= cochlear division of the vestibulocochlear nerve).
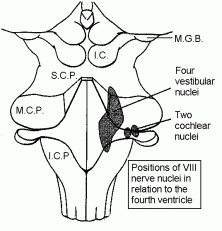
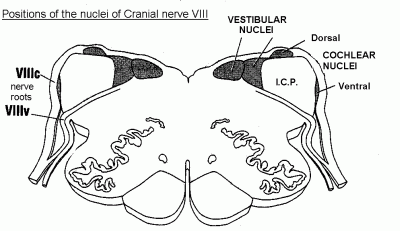
Axons of the cochlear nerve branch as they enter the medulla, and the branches end in the ventral and dorsal cochlear nuclei.
These small images show the locations of the vestibular and cochlear nuclei, and a few other landmarks.
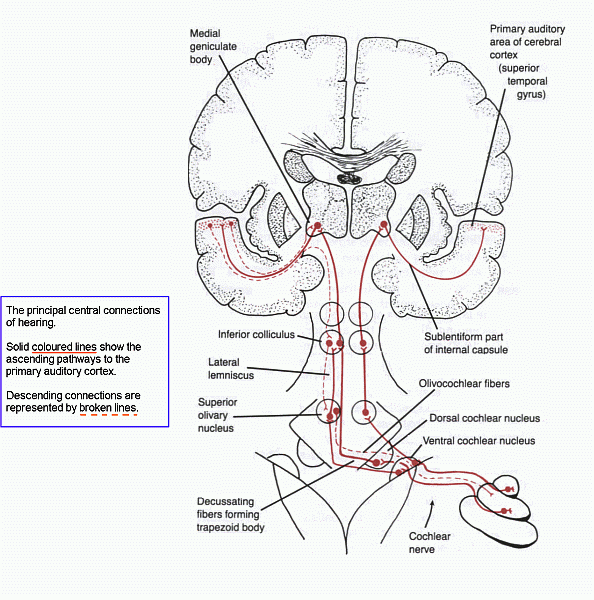
Click here for a diagram of the auditory pathways
Level of transverse section of medulla = rostral end of the olive
Points to note about the central auditory connections.
1. There is more than one ascending pathway, and the number of synaptic interruptions between the organ of Corti and the primary auditory
cortex is variable. Not all the nuclei of the pathways are shown in the diagram.
2. The ascending pathway from each ear is bilateral. Lesions above the level of the cochlear nuclei do not cause
unilateral deafness.
3. Throughout the system there is tonotopic representation:
in each nucleus and tract, cells and fibres are arrayed according to
the frequencies of sound. (This begins in the cochlea, where high
frequencies are detected at the base and low frequencies at the apex.)
4. Each superior olivary nucleus receives input from the left
and right ventral cochlear nuclei. The timing and intensity of signals
from the
two ears are compared, and the resulting output of the superior olivary
nucleus conveys stereophonic
information to the inferior colliculus
and in turn to the thalamus and cerebral cortex. Unilateral damage to
the primary auditory cortex is said to impair recognition of the
sources of
auditory stimuli.
5. The primary auditory cortex (Heschl's convolutions) projects
to the cortex of the posterior part of the superior temporal gyrus.
This is
auditory association cortex, and it interprets the patterns of received signals. In the left hemisphere this is Wernicke's area
for recognizing spoken communication. In the right hemisphere
the corresponding area processes non-verbal sensory input, including
music.
The nearby parts of the parietal lobe (angular and supramarginal gyri)
are also involved in recognizing and responding to complex auditory and
other sensory experiences.
VIII (vestibular). Vestibular system.
|
The vestibular apparatus is concerned with detecting position and movement of the head.
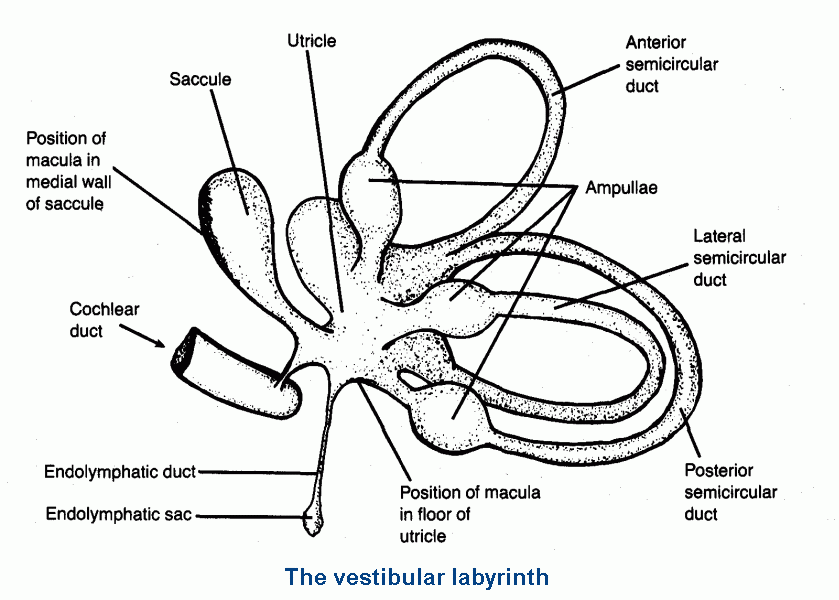
It consists of the saccule, the utricle, and three semicircular ducts.
(Click here for an anatomical picture.
These are parts of a system of tubes and cisterns, the membranous labyrinth, that is entombed in the
petrous part of the temporal bone. The membranous walls are separated from the lining of the bony labyrinth
by perilymph, a fluid similar to CSF. The fluid inside the membranous labyrinth is endolymph, a liquid
similar to intracellular fluid, with a high concentration of K+ and a low concentration of Na+.
The sensory hairs of the receptor cells of the vestibular system (and of the organ of Corti) require an extracellular fluid of this unusual composition in order to work. Endolymph is secreted by the stria vascularis of the cochlear duct, and it is absorbed into the blood from the endolymphatic sac. |
|
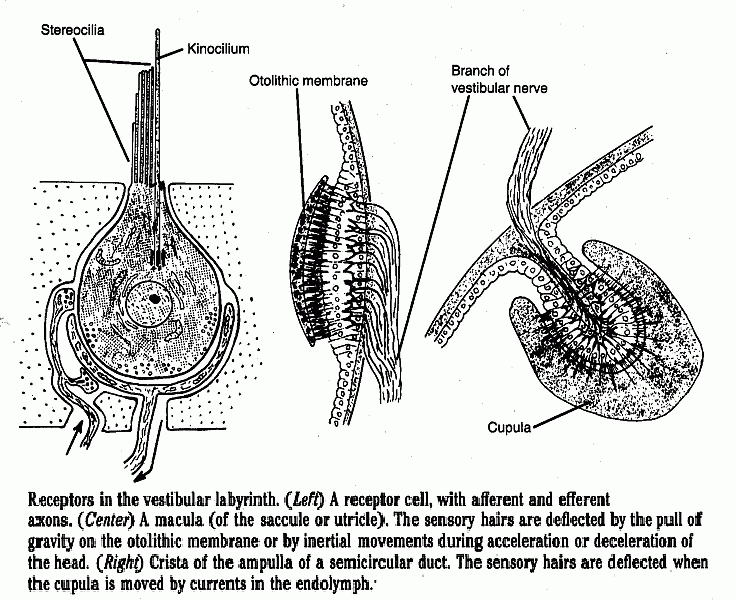
There are vestibular receptors of two types:
| Static labyrinth. Otolithic organs in the utricle and saccule respond to the earth's gravitational force. The otoliths, being denser than endolymph, bend the sensory hairs downward. The direction of bending signals the orientation of the head in space. These receptors also respond to inertial movement of the otoliths, when the body accelerates or decelerates. | Kinetic labyrinth. Receptors in the ampullae of the three semicircular ducts do not have otoliths but the sensory hairs are moved when there is movement (flow) of endolymph relative to the wall of the duct. This occurs with rotational movement of the head *. |
|
Click here for 3 drawings that show the structure of the receptors. |
Clinical testing.
Irrigation of the earhole with either warm or cool water changes
the temperature of the endolymph, which is only
2 or 3 millimetres away from the eardrum. Even a small temperature
change sets up a convection current: warm endolymph flows up, or cold
endolymph flows down. The ampullary receptors move with the flow;
it is as if the head were rotating in the opposite direction.
In a conscious patient this causes vertigo and nystagmus. In a comatose patient the vestibulo-ocular reflex drives both eyes to
look in a direction opposite to the simulated rotation of the head.
This simple clinical examination is called caloric testing. In
its simplest form, it can show that the vestibular system (medulla) is
functionally connected with the ocular motor system (midbrain) in a
comatose patient, providing evidence that the patient is not "brain
dead."
The tract that connects the vestibular nuclei to the oculomotor nuclei is the medial longitudinal fasciculus (MLF).
The MLF also contains fibres that
interconnect the ocular motor (III, IV & VI) nuclei for making
conjugate eye movements, but that's another story.
Central vestibular connections are with the same side of the body and with the same side of the cerebellum.
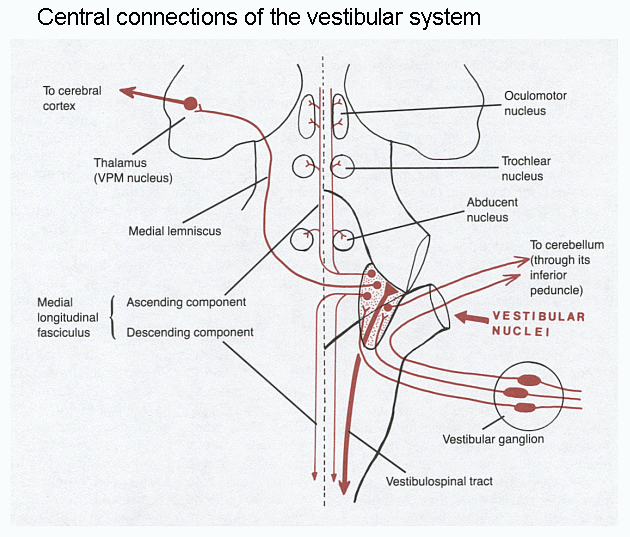
Click here for a larger diagram of the projections.
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
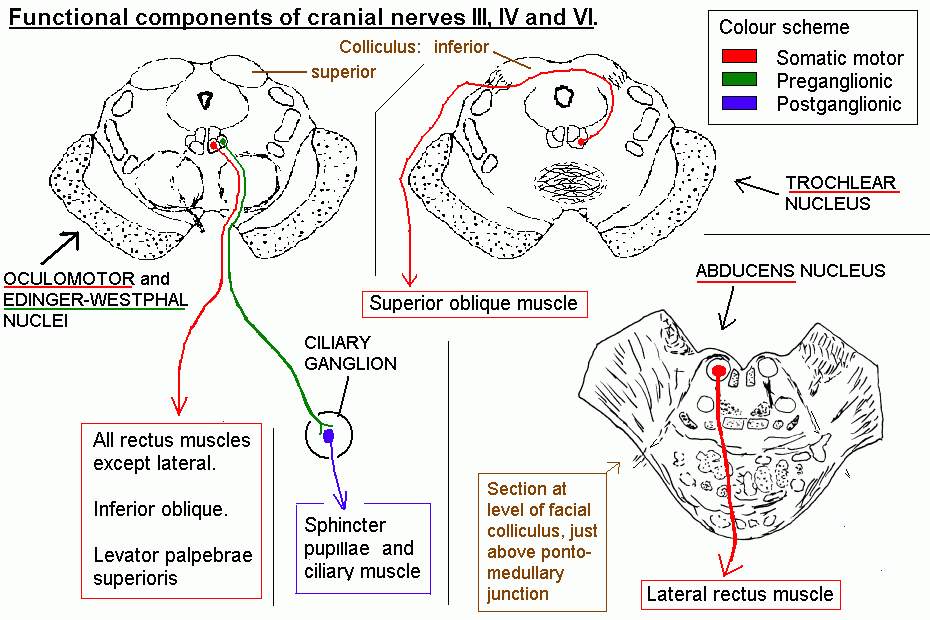
The other cranial nerves: III, IV and VI.
The
three nerves that supply extraocular muscles contain somatic motor
fibres. The oculomotor nerve contains, in addition, preganglionic
parasympathetic fibres.
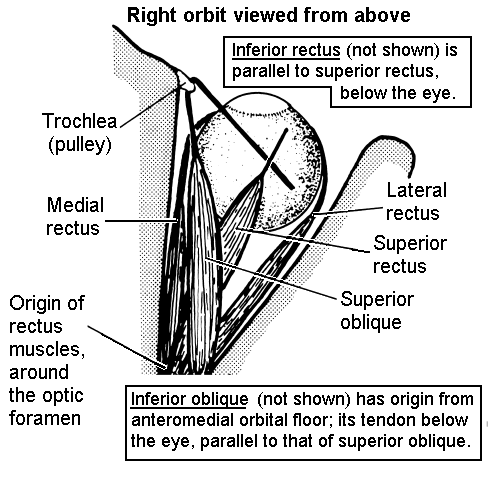
Trochlear nerve supplies only the superior oblique muscle.
Abducens nerve supplies only the lateral rectus muscle.
Oculomotor nerve supplies superior, medial and inferior
rectus muscles, and the inferior oblique. It also supplies the levator
palpebrae superioris (which raises the upper eyelid).
Central connections. ("Deep origins")
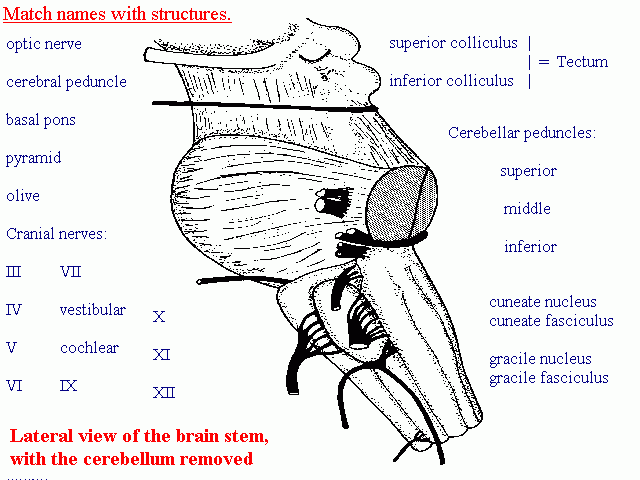
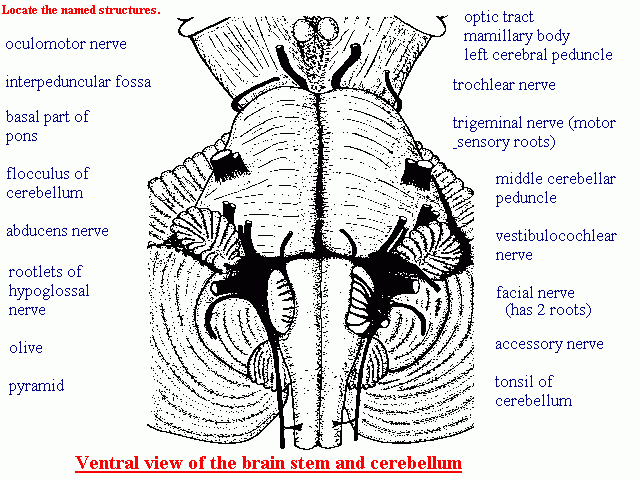
First, remind yourself of the sites of emergence ("superficial origins") of the cranial nerves from the brain stem:
Lateral aspect
Ventral aspect
The nuclei of III, IV and VI near the levels of the emerging nerves.
(Click on the Lateral and Ventral links for larger diagrams.)
The Oculomotor nucleus is a complex of subnuclei, one for
each muscle, located ventral to the periaqueductal grey matter, next to
the midline, at the level of the superior colliculus.
The parasympathetic Edinger-Westphal nucleus is nearby.
Axons course ventrally, passing through the red nucleus.
The most conspicuous feature of a palsy that affects the motor component of III is that the eye is closed, because of paralysis of the levator palpebrae superioris muscle.
| The preganglionic axons from the Edinger-Westphal nucleus end in the ciliary ganglion, which is behind the eyeball. The short ciliary nerves carry unmyelinated postganglionic fibres (axons of the neurons in the ganglion) to innervate smooth muscle: the sphincter pupillae (circular muscle fibres of the iris) and the ciliary muscle, which focuses the lens for near vision. |
| The preganglionic parasympathetic fibres are superficially situated in the nerve and are therefore the first ones to be affected by external pressure. The result is dilation of the pupil, with a sluggish or absent pupillary light reflex. A common cause of third nerve compression is a subdural haematoma pushing downward onto the convexity of a cerebral hemisphere. The uncus is pushed medially and it presses upon and stretches the oculomotor nerve of the same side. |
The trochlear nucleus
is caudal to the oculomotor nucleus, at the level of the superior
colliculus.
The axons of the motor neurons course caudally, dorsally and medially.
They then decussate in the superior medullary velum and leave the
dorsal surface of the brain stem below the inferior colliculus.
Thus, each superior oblique muscle is supplied by the
contralateral trochlear nucleus. The only other muscle that receives
comparable crossed innervation is the superior rectus: axons from the
superior rectus subnucleus of the oculomotor complex cross the midline
and join the third nerve of the opposite side.
A trochlear nerve palsy (due, for example, to diabetic neuropathy, or following a head injury) causes double vision on looking downward. The affected eye is slightly higher than the other (skew deviation), and the patient tilts the head to compensate.
The abducens nucleus
is next to the midline in the floor of the 4th ventricle, at the level
of the ponto-medullary junction. The motor axons pass ventrally.
In addition to motor neurons, the abducens nucleus contains internuclear neurons. These cross the midline, ascend in the contralateral medial longitudinal fasciculus,
and end in the oculomotor nucleus, where they drive the motor neurons
that supply the medial rectus. This arrangement provides for
coordinated horizontal eye movements.
The fibres of the facial nerve loop over the dorsal
aspect of the abducens nucleus. The resulting lump in the floor of the
4th ventricle is called the facial colliculus.
Lesions in this region cause ipsilateral paralysis
of the lateral rectus and facial muscles, and also failure of the
contralateral medial rectus to adduct the eye. A lesion affecting
VI outside the brain stem affects only the lateral rectus.
| Click here for an enlargement of the schematic diagram of the functional components of cranial nerves III, IV and VI. |
|
__________________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
The other cranial nerves: V, VII, IX, X, XI and XII.
In any peripheral nerve there are populations of fibres serving different functions. Each such population is a nerve component. For example, the oculomotor nerve has 2 components: motor and preganglionic autonomic. A nerve in a limb typically contains motor, somatic sensory and postganglionic autonomic components. In some of the cranial nerves there are also components related to the special senses, visceral sensory innervation, and preganglionic parasympathetic fibres (which are also "visceral").
The cranial nerve nuclei in the brain stem deal with the nerve components rather than the nerves.
For example:
First, remind yourself of the sites of emergence ("superficial origins") of the cranial nerves from the brain stem: Lateral aspect Ventral aspect
Trigeminal nerve. This is named for its three large divisions: the ophthalmic, maxillary and mandibular nerves. It has two components:
|
Fibres concerned with touch end in the pontine trigeminal nucleus and in the rostral part of the spinal trigeminal nucleus.
Click here for a diagram of the central connections of the trigeminal nerve.
|
Fibres concerned with pain and temperature turn caudally in the spinal trigeminal tract and end in the caudal part of the spinal trigeminal nucleus. (This is in the caudal half of the medulla and first two cervical segments of the spinal cord, where it blends into the dorsal horn.) |
The neurons for proprioception are exceptional. Their unipolar cell bodies constitute the mesencephalic trigeminal nucleus,
which is lateral to the periaqueductal grey and extends rostrally to
the level of the superior colliculus. The axons of these neurons form
the mesencephalic trigeminal tract and are
then distributed with the maxillary and mandibular divisions of V.
They go to muscle spindles,
receptors in the temporomandibular joint, and pressure receptors around
the roots of the teeth. (This pressure sense is related to
proprioception; reflex connections to the motor trigeminal nucleus
regulate biting and chewing.)
The cells of the mesencephalic trigeminal nucleus are the only primary sensory neurons to have their cell bodies in the central nervous system. |
Facial nerve. This supplies the facial muscles and also has sensory and preganglionic autonomic components. The primary sensory neurons have their cell bodies in the geniculate ganglion. The functional components of VII are distributed in different branches of the nerve.
| Click here for an enlarged diagram of the components of the facial nerve. |
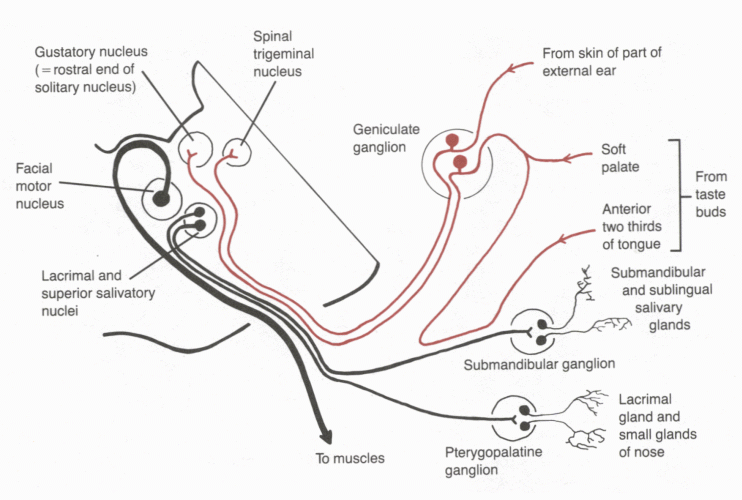
|
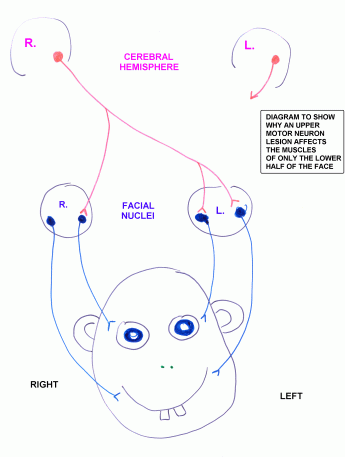 When a facial paralysis is due to a lesion of the facial motor nucleus or of the nerve (a lower motor neuron lesion)
all the muscles of the same side of the face are affected. (There may
also be inadequate lacrimal or salivary secretion, or a localized taste
deficit, depending on the exact location of the lesion.)
An upper motor neuron lesion affects only the lower half of the contralateral side of the face, for the reason explained in
this diagram.
When a facial paralysis is due to a lesion of the facial motor nucleus or of the nerve (a lower motor neuron lesion)
all the muscles of the same side of the face are affected. (There may
also be inadequate lacrimal or salivary secretion, or a localized taste
deficit, depending on the exact location of the lesion.)
An upper motor neuron lesion affects only the lower half of the contralateral side of the face, for the reason explained in
this diagram.
Glossopharyngeal nerve. This too has motor, sensory and preganglionic components. Nuclei are shared with other cranial nerves, notably X.
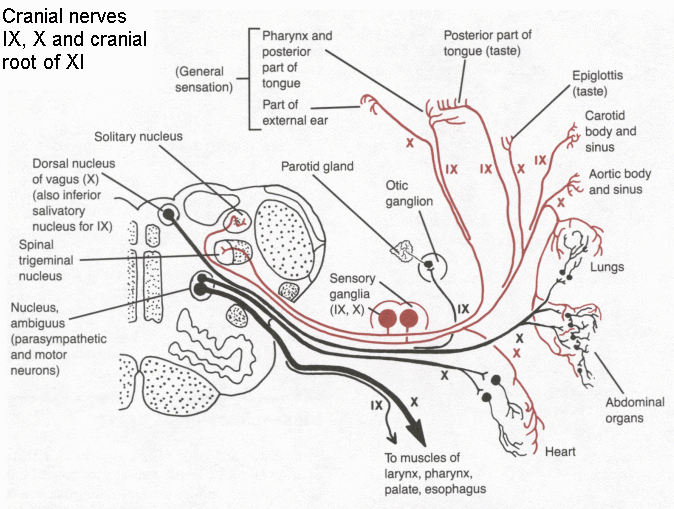
Vagus nerve. This nerve, like IX, has five components and two sensory ganglia. Its branches extend beyond the head and neck into the thorax and abdomen, hence the name (Latin vagus = wandering).
Click here for a diagram showing the central connections of the glossopharyngeal and vagus nerves.
Accessory nerve. This is formed intracranially by the union of two roots.
(Click here to see a drawing of all the cranial nerve roots.)
All the fibres in both roots are motor. The spinal root contains axons of motor neurons that form a cell column, the accessory nucleus
in the first 5 cervical segments of the spinal cord. The spinal root
ascends in the subarachnoid space, enters the cranial cavity by way of
the foramen magnum, and fuses with the cranial root.
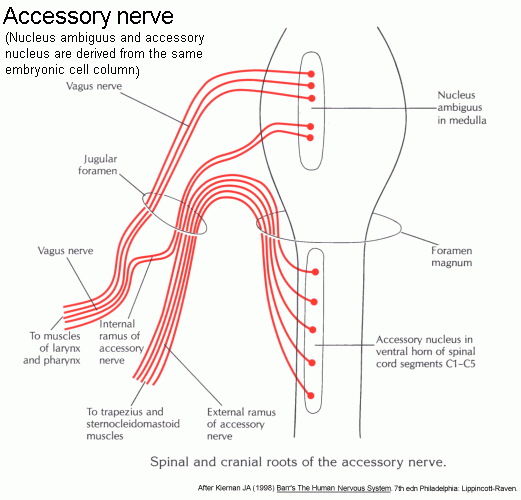 The cranial root consists of the axons of some of the cells in the nucleus ambiguus, notably neurons that supply laryngeal muscles.
The cranial root consists of the axons of some of the cells in the nucleus ambiguus, notably neurons that supply laryngeal muscles.
After fusion of the roots, the accessory nerve leaves the cranial
cavity by way of the jugular foramen, alongside cranial nerves IX and
X. The axons from the nucleus ambiguus then leave XI in its small
internal ramus, which anastomoses with the vagus nerve. The larger
external ramus of XI continues into the posterior triangle of the neck
and supplies motor fibres to the trapezius and sternocleidomastoid muscles.
(Click here for enlarged diagram of the components and roots of IX and its association with X.)
Disorders affecting IX, X and XI. Damage to the motor
neurons in the nucleus ambiguus or to their axons in branches of the
nerves causes hoarseness (larynx) and unsymmetrical elevation of the
soft palate (uvula deviates to the normal side). If the accessory nerve
is injured in the neck the shoulder on the same side is lower
(trapezius) and the head is turned towards the same side by the action
of the contralateral sternocleidomastoid muscle.
Upper motor neuron lesions do not affect the larynx and pharynx
because the nucleus ambiguus receives bilateral afferents from the
cerebral hemispheres. A hemispheric lesion typically causes weakness of
the contralateral trapezius and the ipsilateral sternocleidomastoid -
muscles that both move the face in the same direction.
Although IX is the conduit for much pain and discomfort in diseases of
the throat and middle ear, sensory deficits in the territory of IX, X
and XI are seldom encountered.
Disorders of parasympathetic functions of IX and X are also
unusual. Surgical transection of both vagus nerves at the level of the
diaphragm (vagotomy) suppresses gastric acid secretion and inhibits
opening of the pyloric sphincter.
Hypoglossal nerve. This is another nerve with a single, somatic motor component.
In a lower motor neuron hypoglossal palsy the
affected side of the tongue shrinks (denervation atrophy) and deviates
to the paralysed side when protruded. In an upper motor neuron lesion
(such as a hemispheric stroke) the tongue deviates to the weakened
(contralateral) side but does not atrophy. There is partial bilateral
descending control of the hypoglossal nucleus, so the effects of a
upper motor neuron weakness are not severe and there is recovery.
The cell bodies are in the hypoglossal nucleus,
which is next to the midline in the caudal half of the floor of the 4th
ventricle and extending caudally into the rostral part of the closed
medulla. The axons innervate the extrinsic and intrinsic muscles of the
tongue.
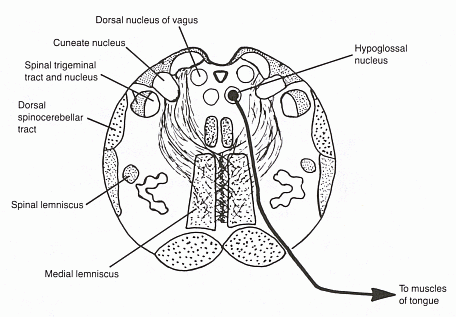 Click here for a larger diagram.
Click here for a larger diagram.
How much of all this cranial nerve stuff do I really need to know for 350a? The only way to come to grips with the cranial nerves is to know the components and the associated nuclei, ganglia and functions. For a neuroanatomy course you are not expected to know the names and topographical details of the branches in the head and neck (though in the case of the facial nerve this information facilitates learning the functional components). You should know the neuroanatomical reasons for clinical abnormalities.
A useful exercise to test your knowledge is the following:
Set up a table with the functional components as the columns and the
numbered cranial nerves as the rows. In each cell of the table write
the names of nuclei, ganglia and functions. For example:
(Only a few cells have been filled in above, as examples. A column could be added for general visceral sensation, and the table could have rows for cranial nerves I, II and VIII.)
Component:
Nerve
Motor
Somatic sensory
Special sensory
Preganglionic
III
Oculomotor nucleus, eye muscles except lat. rect. & sup. obl.
Edinger Westphal nucleus, ciliary ganglion, focusing & pupil constriction
IV
VI
V
Motor trig. nuc. Masticatory & related muscles
Face etc; 3 divisions; trigeminal ganglion, pontine & spinal V nuclei. Pain & temp. to spinal nucleus
VII
IX
Nucleus ambiguus, stylopharyngeus only
IX ganglia, pharynx, back of tongue, middle ear
Taste, back of tongue
X
Nucleus ambiguus: cardiac ganglia. Dorsal nucleus of X: intrinsic neurons of stomach etc.
XI
XII
__________________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
Hypothalamus and pituitary gland.
[ For comfortable viewing of this section, your web browser window should occupy most of the width of the screen ]
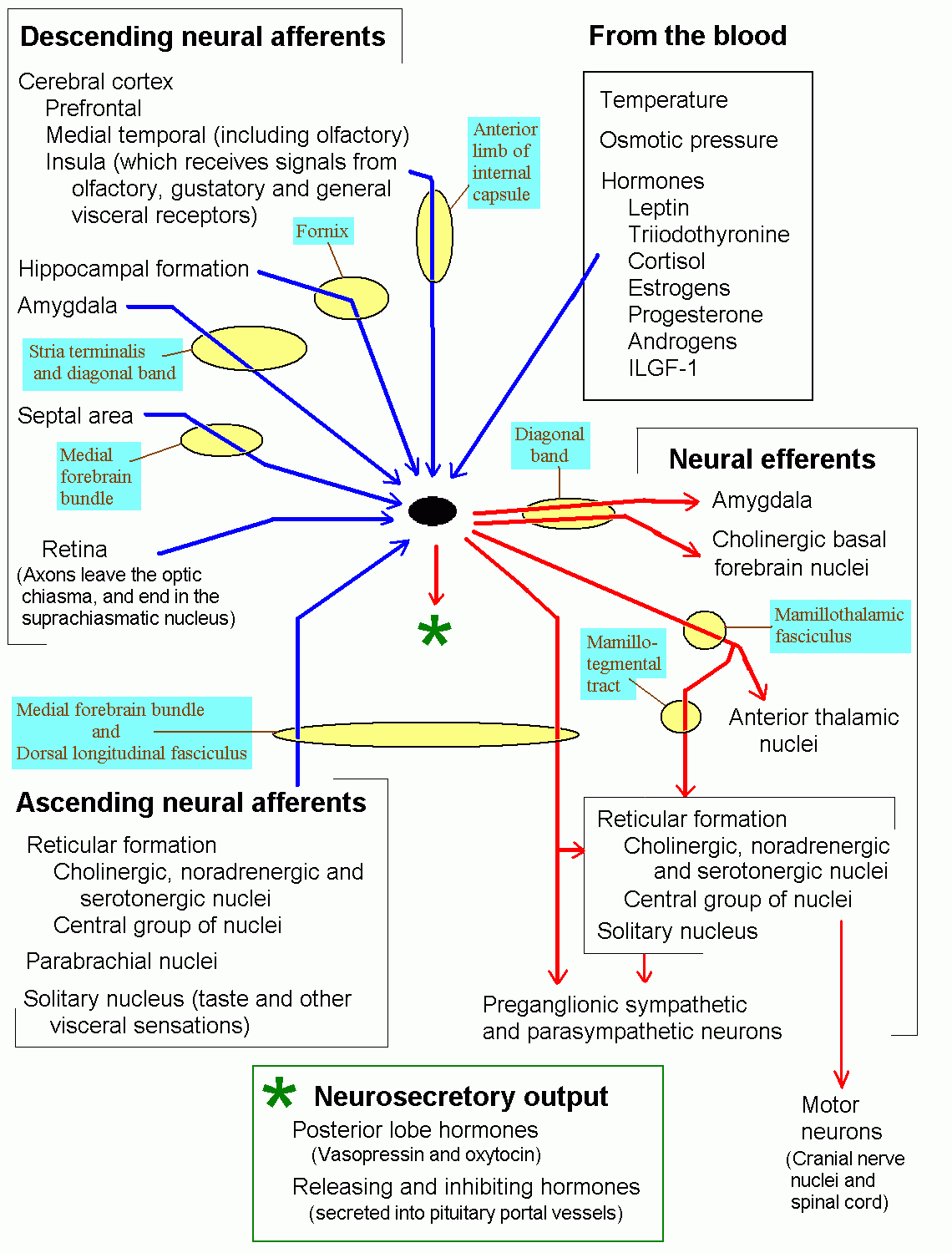
Click here to download a diagram of hypothalamic
connections and functions. It's quite complicated
but might be helpful for revision.

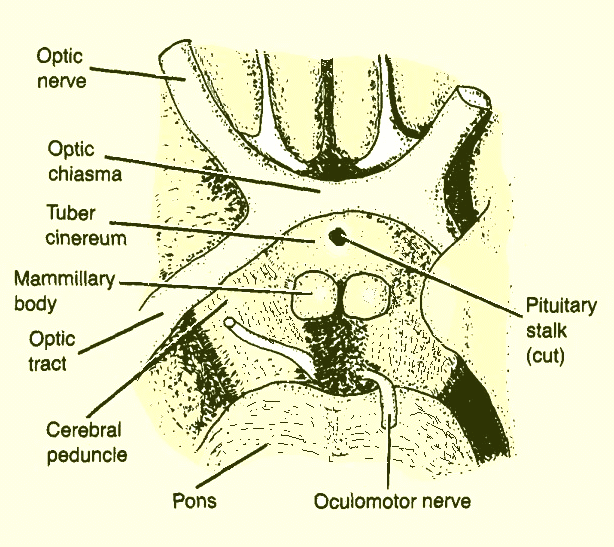
Anatomical landmarks of the hypothalamus.
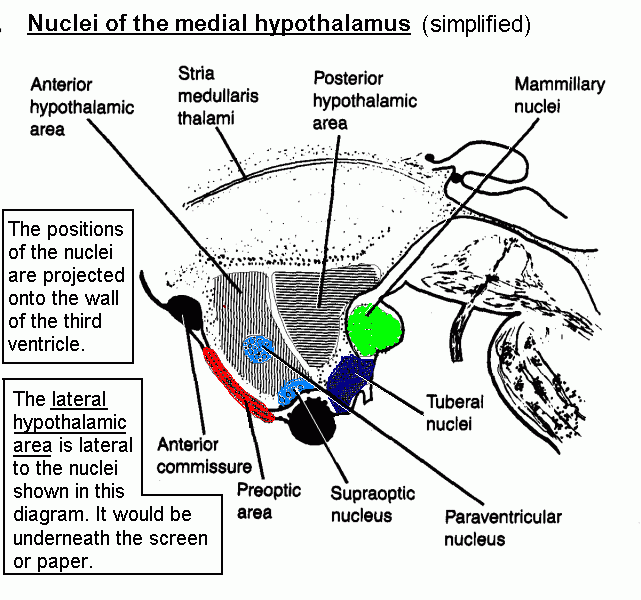
(Hypothalamic nuclei are projected onto a mid-sagittal section.)
Some neural connections of the hypothalamus.
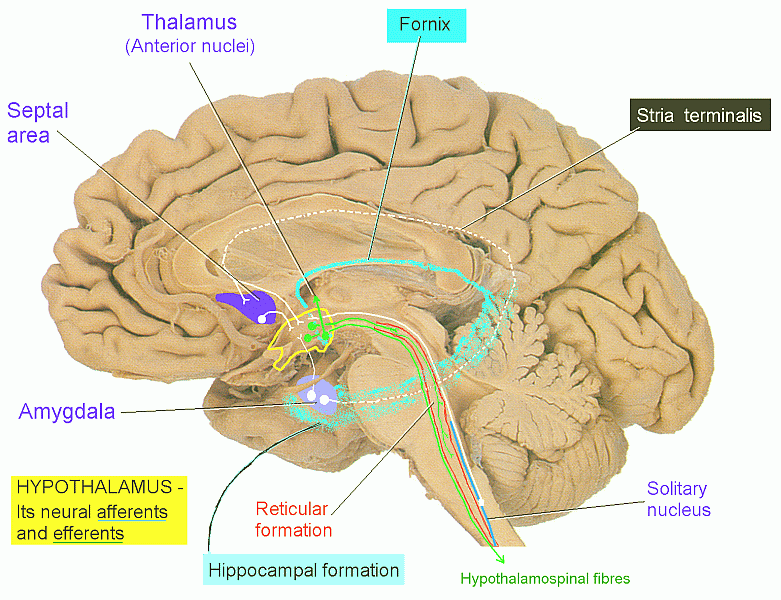
Some hypothalamic neurons project directly to preganglionic neurons of the autonomic nervous system.
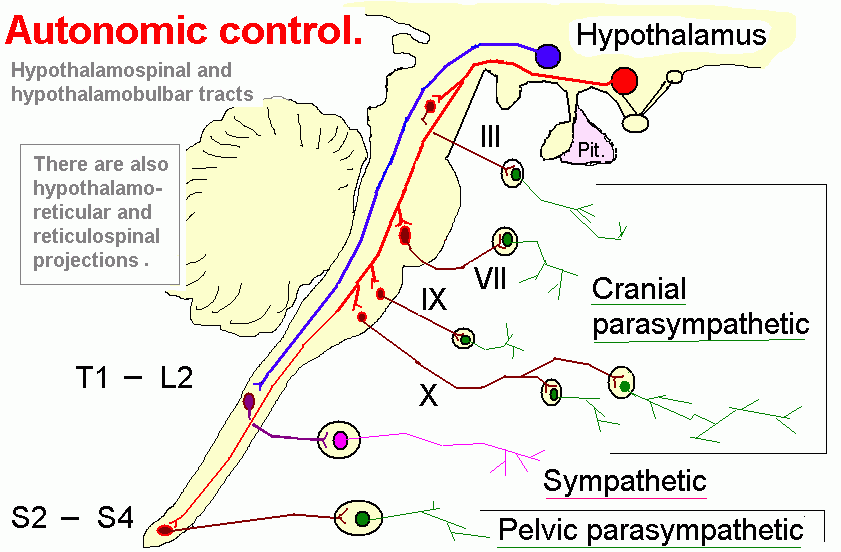
Regulation of body temperature is an important activity controlled by the hypothalamus.
 Heat is produced by contracting muscles. The pathway is probably from the posterior hypothalamus to parts of the pontine
and medullary reticular formation that give rise to motor reticulospinal fibres. Shivering is one way of raising a low body temperature.
Heat is produced by contracting muscles. The pathway is probably from the posterior hypothalamus to parts of the pontine
and medullary reticular formation that give rise to motor reticulospinal fibres. Shivering is one way of raising a low body temperature.



The other hormone of the posterior lobe is oxytocin, named for its action of causing the uterus to contract.
Oxytocin also causes milk ejection, through a reflex initiated by suckling. The myoepithelial cells of the mammary gland contract, pushing the milk from the secretory units into the duct system. The circulating oxytocin also makes the uterus contract - an important event of maternal recovery following birth.
The hypothalamus, as well as the reticular formation of the brain stem, is involved in the sleep/waking cycle.

The mamillary bodies are part of the circuitry of the hippocampal formation.

The hypophysial (= pituitary) portal veins deliver high concentrations of hypothalamic releasing hormones to the anterior lobe of the pituitary gland.
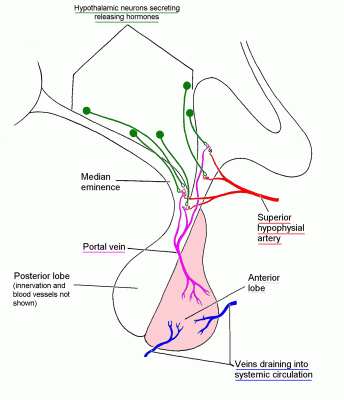
(There are also hypothalamic release inhibiting hormones.)
The hypothalamic releasing hormones have names that end in -RH. For
example, CRH for corticotrophin releasing hormone, and LHRH for
luteinizing hormone releasing hormone.
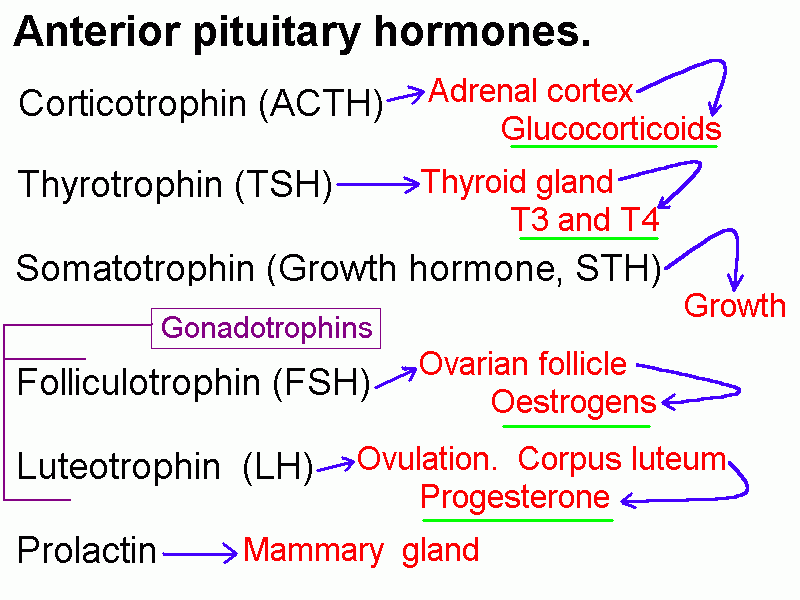
Secretion of prolactin is controlled mainly by an inhibiting hormone, which is dopamine. Growth hormone secretion is controlled by GHRH and also by a release inhibiting peptide called somatostatin.
Benign tumours of the anterior lobe are quite common. Most do not produce hormones. Rarely a tumour secretes excessive amounts of a hormone. For example, an adenoma that produces growth hormone will cause gigantism in a child or acromegaly in an adult. Pituitary tumours enlarge in a rostral direction and presss on the optic chiasma, causing bitemporal hemianopia.
__________________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
Functional neuroanatomy of the prefrontal cortex, amygdala, and hippocampal formation. Disorders of the frontal and temporal lobes.
The prefrontal cortex (= prefrontal lobe) is the frontal cortex anterior to the areas specifically involved in motor control (including eye movements and speech). It includes the frontal pole and the orbital surface of the frontal lobe.
Diagram of connections of prefrontal cortex
Lesions that damage the prefrontal cortex bilaterally
(or sever its connections with subcortical structures) cause a syndrome
in which there is a profound personality change, with loss of the
ability to predict consequences of actions or to adapt behaviour based
on experience. The syndrome is attributable especially to loss of
function of the orbital cortex and the cortex of the frontal pole.
Other anterior frontal lesions can cause difficulty placing objects or
ideas in correct sequences.
Causes of prefrontal lobe dysfunction include trauma,
surgery (prefrontal leukotomy), ischaemia, syphilis and Pick's disease.
The amygdala (= amygdaloid body) is inside the temporal pole - internal to the uncus, on the end of the tail of the caudate nucleus, and bulging into the temporal horn of the lateral ventricle. In a coronal section, the amygdala is seen as a mass of grey matter continuous with the cortex in the region of the uncus. The name amygdala is Latin for almond.
Diagram of connections of amygdala
Electrical stimulation of the human amygdala
causes feelings of fear and sometimes also of anger. Increased neuronal
activity can be demonstrated (by fMRI) in subjects shown emotionally
evokative pictures.
Destructive lesions in the human amygdala are associated
with damage to the nearby hippocampal formation and visual association
cortex of the temporal lobe.
The hippocampal formation is (for Anatomy 350a purposes) the same thing as the hippocampus as seen in dissected specimens.
The name hippocampus is Latin for sea-horse, probably applied to the appearance seen in a coronal section of the temporal lobe.
The hippocampus and dentate gyrus constitute a body of cerebral cortex with only one layer of neuronal cell bodies. In the subiculum the hippocampus merges with the 3-layered cortex of the entorhinal area, which is the anterior end of the parahippocampal gyrus. The name "entorhinal" means it's medial to the rhinal sulcus,
which, in the human brain, is the anterior end of the collateral sulcus.
In rodents, rabbits and lowly mammals
that eat worms and insects, the rhinal sulcus is the only visible
indentation on the surface of the cerebrum, and the entorhinal area can
account for one third or more of the cerebral cortex. The human
entorhinal area (2 or 3 square centimetres) is much bigger than the
equivalent area in little furry animals. Nevertheless, the entorhinal
area and hippocampus are used by squirrels to survive in winter, by
finding seeds and nuts that they have hidden.
The connections of the hippocampus and the evidence for its involvement in memory were briefly reviewed in the
notes for the previous lecture.
__________________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
Neuroanatomy of language and memory.
Click here for a diagram showing association and language areas
Sensory association areas.
The primary sensory areas are connected by subcortical association fibers with the appropriate sensory association areas.
A primary sensory area receives information about the events reported by an ascending pathway, but it cannot recognize or appreciate the importance of the sensation. The higher functions of recognition and use of sensory data reside in areas of association cortex. These large areas, not always clearly defined, are adjacent to the primary sensory areas.
Thus the association cortex for general somatic sensation is posterior to the postcentral gyrus.
The association area for vision occupies the occipital lobe,
together with the adjacent posterior parietal cortex and the
inferolateral cortex of the temporal lobe.
The auditory association cortex is posterior to the primary area on the superior and lateral surfaces of the superior temporal gyrus.
The olfactory association area is the entorhinal cortex and
the more posterior parts of the parahippocampal gyrus, together with
part of the orbital cortex of the frontal lobe.
In addition to association fibers from the primary sensory areas, the sensory association areas receive projection fibers from several thalamic nuclei.
A large lesion in an association area causes difficulty with recognition of things that are heard, seen or felt. Disorders of the sensory association areas cause various kinds of agnosia. [Recognition of significance is gnosis (Greek for "knowledge").]
An agnosia may be visual, auditory, or tactile. The most common form of tactile agnosia is astereognosis, the inability to identify objects felt with the hand. The word astereognosis is also commonly used for inability to discern differences in shape and texture with the fingers. This symptom is usually due to a lesion in the contralateral parietal lobe, posterior to the hand area of the primary somaesthetic cortex.
Click here for diagram of somatic sensory areas and lesions that cause agnosias.
Language areas.
Cortical areas concerned with verbal and written communication, are in the temporal, parietal, and frontal lobes. These areas are concerned with language in only one of the hemispheres, usually the left.
Click here for diagrams of the cortical language areas and of their interconnections.
The temporoparietal area is the receptive language area, which includes parts of the superior temporal, angular, and supramarginal gyri. Its integrity is necessary for the recognition of spoken and written words. The anterior part of this cortical region, which is the auditory association cortex, is called Wernicke's area, after Carl Wernicke (1848-1905), a German clinical neurologist. The receptive language area receives afferent fibers from the auditory and visual association areas of both hemispheres. It is also connected with the expressive language area in the left frontal lobe.
The expressive language area is often called Broca's area. It is in the frontal lobe, anterior to the lower end of the precentral gyrus.
Paul Broca (1824-1880), a French physician,
was one of the first people to detect functional differences between
the left and right cerebral hemispheres.
The integrity of Broca's area is necessary for producing speech to communicate ideas formulated in the mind.
The supplementary motor area is also essential for speech.
Disordered speech due to loss of function of cortical language areas is called dysphasia, or aphasia if the condition is very severe. These works have Greek roots meaning difficulty with or loss of speech.
Apraxia
Apraxia is clumsiness of movement due to cerebral cortical dysfunction. A typical cause is an infarct in the somatosensory association cortex in the medial part of the superior surface of the parietal lobe. This normally feeds signals related to proprioception to the motor cortex. Without properly processed proprioceptive input. Inaccurate instructions to the pontine nuclei, motor neurons etc result in clumsy movements of the contralateral hand. Which artery supplies the medial part of the superior surface of the parietal lobe?
Click here for diagrams of the motor cortical areas and of locations of lesions that cause apraxia etc.
Memory.
It
has long been generally agreed that memory for perceived facts and
events is a function of the forebrain, and especially of the cerebral
cortex.
If the word "memory" is more
liberally interpreted, then it is a property of all parts of the brain,
because neuronal activity everywhere is modified by earlier neuronal
activity. It is possible to observe conditioned responses even in
certain types of invertebrate animal that do not have anything
equivalent to a brain. Memory can be displayed by cells that are not
neurons, too. For example, the immune system responds more rapidly and
intensely to previously encountered antigens than to antigens presented
for the first time - the basis of immunization against infectious
diseases, and the value of booster shots.
Experiments by Lashley (American psychologist) in the 1920s provided evidence against the involvement of specific parts of the forebrain in memory. He removed numerous parts of dogs' cerebral hemispheres and measured the resulting impairment of learned behaviour. The severity of the impairment increased with the amount of removed cerebral cortex, but was not related to which areas of cortex were removed. Lashley's conclusion: Learning, storage and recall occur diffusely throughout the cortex.
In 1937 Kluver & Bucy (American neuropathologist & neurosurgeon) published a classic paper describing the effects of bilateral removal of the the temporal lobes in monkeys. Among other abnormalities, the animals lost the ability to learn new tricks. (This is just part of the Kluver-Bucy syndrome.) Later work, including studies of human subjects with bilateral temporal lobe lesions (from surgery and disease) indicated that the hippocampal formation was the part of the temporal lobe involved in learning. Through association fibres, the hippocampus can receive input from all regions of the cerebral cortex. The commonest disease that impairs recent memory is Alzheimer's - in which the earliest degenerative changes are in the entorhinal area and hippocampus. Memory deficits result also from bilateral lesions that interrupt the connections of the hippocampal formation.
The
evidence for the hippocampi as parts of the brain essential for
acquisition of memories is currently widely accepted, but it is also
challenged by some greatly respected investigators. The challengers
point out that bilateral hippocampal lesions are large and are always
accompanied by damage to other parts of the cerebral hemisphere.
Lashley might be right.
The postulated role of the hippocampus is in acquisition and short-term storage of remembered experiences. Long-term storage is not affected by bilateral hippocampal lesions or in the early stages of Alzheimer's disease. Procedural memory (for learned motor skills) resembles long-term memory in this respect.
Click here for two diagrams of hippocampal circuitry
Against Lashley's notion of disseminated memory storage:
___________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
Examples of clinical diagnosis by application of neuroanatomical knowledge:
The consequences of an arterial blockage illustrate both the distribution of the artery and the functions of the structures it supplies. By the end of a course in functional human neuroanatomy students should be able predict the effects of localized lesions and should be able to interpret the clinical symptoms and signs to determine the location of a lesion and, in typical examples, the identity of the obstructed artery.
| (a) Cerebral artery occlusions. |
Click here for a map of the cortical areas supplied by the three cerebral arteries.
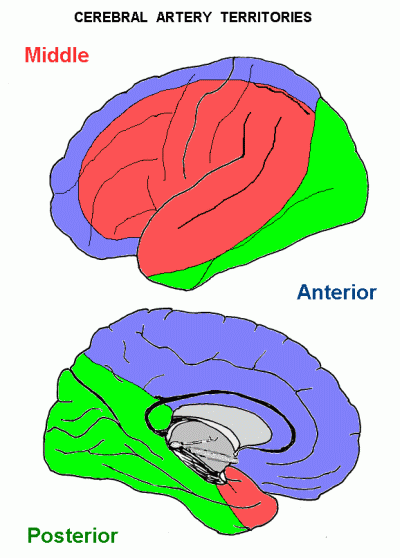
|
Internal carotid artery.
This
vessel is frequently obstructed by atheroma, which commonly gives rise
to transient symptoms of cerebral ischaemia known as transient ischaemic attacks (TIAs). Complete, sudden obstruction cuts off most of the blood flow in the anterior and middle cerebral arteries.
Contralateral hemiplegia with sensory deficits. If on the left side (in 90% of people), severe aphasia.
There may also be blindness in the
ipsilateral eye (ophthalmic artery) with loss of the temporal visual
field of the contralateral eye (anterior choroidal artery supplies
optic tract).
If the internal carotid artery is occluded after a prolonged period of
progressive narrowing by atheroma, the anastomotic vessels at the base
of the brain enlarge, so that the cerebral hemisphere receives blood
from the contralateral side and from the vertebro-basilar system.
Ischaemia may then be restricted to regions supplied by certain
branches that do not receive adequate collateral perfusion. For
example, a gradual internal carotid occlusion might cause infarction in
the territory of anterolateral central branches of the middle cerebral
artery that supply the posterior limb of the internal capsule.
Middle cerebral artery.
Occlusion
of the vessel at its origin causes contralateral upper-motor-neuron
paralysis of the upper limb and face, with global aphasia if
left-sided. Lower limb functions are spared. Infarction of the
geniculocalcarine tract, deep to the parietal cortex, results in
contralateral hemianopia.
Obstruction of branches of the MCA results in fragments of the complete syndrome, such as monoplegia, upper motor neuron facial paralysis, or receptive aphasia.
Anterior cerebral artery.
Paralysis and sensory deficits in the contralateral lower limb and perineum. Urinary incontinence.
A proximal lesion - obstructing
the origin of the anterior cerebral artery - causes hemiplegia
affecting the limbs and lower face, because an early branch, the
recurrent artery of Heubner, supplies the internal capsule. Proximal
lesions may also cause ipsilateral anosmia from infarction of the
olfactory bulb and tract.
Posterior cerebral artery.
Infarction of the occipital lobe causes contralateral hemianopia. Damage to the hippocampal formation results in a memory deficit, but this soon recovers. Bilateral posterior cerebral artery occlusions can result in a permanent inability to form new memories, in addition to blindness in both eyes.
__________________________________
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
Examples of clinical diagnosis by application of neuroanatomical knowledge:
(b) Brain stem strokes.
The vertebral and basilar arteries and their branches may be obstructed by atheroma or embolism. Often blockage of a larger vessel causes ischaemia only in the territory of a smaller branch. For example, the syndrome of occlusion of the posterior inferior cerebellar artery is often due to an obstructed vertebral artery.
Basilar artery thrombosis. Coma and decerebrate rigidity, soon followed by death from respiratory failure.
(A large pontine haemorrhage results in sudden death.)
Basilar artery embolism. The embolus typically lodges at the terminal bifurcation of the basilar artery and obstructs the posterior cerebral and superior cerebellar arteries, including the central branches from the proximal part of the posterior cerebrals. Coma (from infarction of reticular formation of midbrain and rostral pons); diverging eyes with fixed, dilated pupils (bilateral infarction of fibres of III). In less severe cases there is recovery of consciousness, and the residual top of the basilar syndrome includes the oculomotor paralysis, together blindness or lesser visual field defects (occipital lobes) and disturbances of memory or behaviour (temporal lobes).
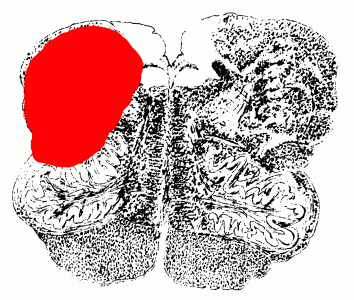 Posterior inferior cerebellar artery (PICA). (Wallenberg's syndrome) This is the commonest brain stem stroke.
There is loss of pain and temperature sensation from the same side of the face (spinal trigeminal tract and nucleus) and from the contralateral
limbs (spinothalamic tract); ipsilateral paralysis of vocal cord and
palatal muscles (nucleus ambiguus); ipsilateral ataxia (inferior
cerebellar peduncle), vertigo (vestibular nuclei), and an ipsilateral
Horner's syndrome (due to interruption of hypothalamospinal or
reticulospinal fibres destined for the preganglionic sympathetic
neurons in the upper thoracic segments of the spinal cord).
[ This is a reproduction of Wallenberg's original (1908) drawing. ]
Posterior inferior cerebellar artery (PICA). (Wallenberg's syndrome) This is the commonest brain stem stroke.
There is loss of pain and temperature sensation from the same side of the face (spinal trigeminal tract and nucleus) and from the contralateral
limbs (spinothalamic tract); ipsilateral paralysis of vocal cord and
palatal muscles (nucleus ambiguus); ipsilateral ataxia (inferior
cerebellar peduncle), vertigo (vestibular nuclei), and an ipsilateral
Horner's syndrome (due to interruption of hypothalamospinal or
reticulospinal fibres destined for the preganglionic sympathetic
neurons in the upper thoracic segments of the spinal cord).
[ This is a reproduction of Wallenberg's original (1908) drawing. ]
Click here for a photo of a
Weigert-stained section of the medulla with annotations explaining the
clinical features of a lateral medullary lesion.
Horner's syndrome, due to loss of sympathetic action on the eye, comprises miosis (small pupil), ptosis (drooping eyelid, but not closed as in a IIIrd nerve palsy) and apparent enophthalmos. (The eyeball appears further back in the orbit than the normal eye; this appearance is caused by the ptosis.) Johann Friedrich Horner (1831-1886) was a Swiss ophthalmologist. His syndrome can have several different causes.
Anterior inferior cerebellar artery (AICA). Ipsilateral ataxia (cerebellum, middle cerebellar peduncle); vertigo and ipsilateral deafness (from inner ear infarction, the labyrinthine artery being in most people a branch of AICA).
Other brain stem syndromes.
Do not attempt to memorize the six examples that
follow. Except in the case of Parinaud's syndrome, you should be able
to deduce the disordered function from the structures damaged by the
lesion. These are classical syndromes that have contributed to our
understanding of the functional neuroanatomy of the brain stem.
For Anatomy 530a you should be
sufficiently familiar with sections of the midbrain to recognize the
consequences of the lesions causing the syndromes of Weber and
Benedikt. You do not need to be visually familiar with sections of the
pons, but you are expected to know that the nuclei of VI and VII are
dorsally located at a caudal
level of the pons, and that descending motor fibres (corticospinal etc)
pass through the ventral part of the pons. You must also understand the
differences between upper and lower motor neuron lesions that affect
the facial and other muscles.
Pontine lesions that impair conjugate eye movements
are important for clinical diagnosis and functional neuroanatomy, but
there will be no specialized neuro-ophthalmological questions in the
final exam.
Midbrain lesions
| Weber's syndrome | Benedikt's syndrome | Parinaud's syndrome |
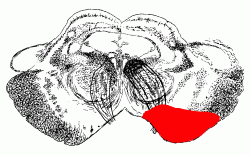
|
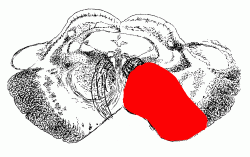 |
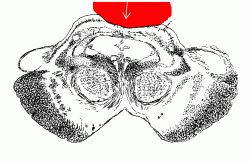
|
|
Fibres of III.
Descending motor fibres in cerebral peduncle. |
Fibres of III.
Descending motor fibres in cerebral peduncle. Red nucleus and nearby cerebellothalamic fibres. |
Pressure on superior colliculi, usually by a pineal tumour, is transmitted to pre-oculomotor nuclei in region of posterior commissure and periaqueductal region |
|
Ipsilateral oculomotor palsy.
Contralateral hemiplegia. |
Ipsilateral oculomotor palsy.
Contralateral hemiplegia. Contralateral tremor. (Tremor is usually attributed to interruption of cerebellothalamic fibres but it may be continuous, from involvement of pallidothalamic fibres rostral to the red nucleus. |
Paralysis of upward gaze and convergence, often accompanied by other pupillary and eye movement abnormalities.
Various pre-oculomotor nuclei, concerned with vertical eye movements, are present in the affected region. |
Pontine lesions
| Millard-Gubler syndrome | Raymond's syndrome | Foville's syndrome | |
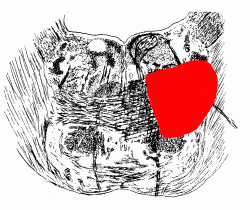
|
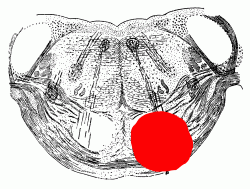
|
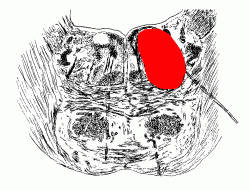
|
|
|
Facial motor nucleus and fibres of VII.
Descending motor fibres (but most are ventral to the lesion). |
Descending motor fibres.
Fibres of abducens nerve. |
Abducens nucleus.
Facial motor nucleus. |
|
|
Ipsilateral lower motor neuron facial paralysis.
Contralateral upper motor weakness of limbs (which recovers with time). |
Contralateral hemiplegia.
Ipsilateral VI nerve palsy (but no abnormality of movements of contralateral eye). |
Ipsilateral VI nerve palsy and inability of the
contralateral eye to adduct when attempting a conjugate movement. The
contralateral eye does adduct with convergence to look at a near
object.
Ipsilateral lower motor neuron facial paralysis. |
|
Accounts of these syndromes were published between 1855 and 1895. The earlier investigators such as Millard and Foville had little understanding of the neuroanatomy. In the later studies, such as those of Raymond and Wallenberg, great efforts were made to fit the clinical findings to tracts and cranial nerve nuclei. The notion of the neuron as a cell and physiological unit, in its infancy in 1895, was finally proved in the 1950s. The notion of chains of neurons, linked by synapses, resulted from the work of Sir Charles Sherrington and other neurophysiologists who worked in the first half of the 20th century.
Acoustic neuroma. The effects of this benign
tumour are presented in the lecture as a diagnostic exercise. The glial
cells (Schwann cells) of the vestibular nerve proliferate and the
resulting mass presses on the cochlear nerve, causing deafness. This
happens where the two divisions of cranial nerve VIII are side by side
and surrounded by bone, in the internal auditory meatus. The facial
nerve is also present in this bony canal, and it is compressed.
The tumour expands out of the internal meatus into the subarachnoid
space, in the angle formed by the pons, medulla and cerebellum. As the
tumour gets bigger it hits the side of the medulla (spinal trigeminal
tract), and expands upward (roots of V) and downward (rootlets of IX,
X).
Click here to view a graphics file
(acnroma2.gif) that relates the clinical features of an acoustic
neuroma to the cranial nerves involved.
Test yourself. Why does this tumour, expanding
into the pontocerebellar angle, spare cranial nerves VI and XII?
What structures other than cranial nerve roots might be pressed
upon by a tumour in the pontocerebellar angle? (The tumour is benign
only in the pathological sense of not being malignant. It will kill the
patient if it goes on growing. How?)
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
-----------------------------
Last updated: 13th November 2007
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
These
multiple choice and short-answer questions test the subject in greater
detail than is currently required for Anatomy 530a.
Don't expect to be able to answer all of them!
Neuro Quiz. A collection of about 300 questions, grouped by subject. Answer by typing a word or short
phrase. The correct answer is then shown and explained.
All the files from the NEUROQUIZ.ZIP archive must reside in the same directory.
The command BQ starts the
program. For more information read the text file BQ.DOC
Neuro-MCQ. About 140 multiple choice questions in Neuroanatomy. They can be chosen in groups by subject
or at random from the whole collection.
A single file, NMCQ.EXE, is all that is needed to run this program.
Return to "Instructional resources."
Return to beginning of Anatomy 530a home page
Return to Anatomy 530a list of topics
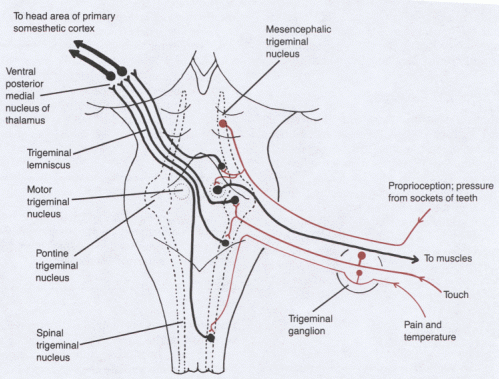
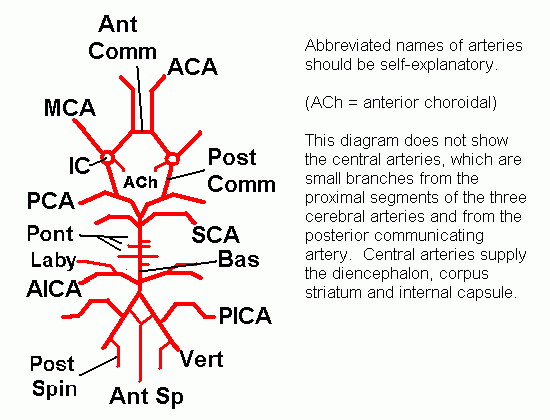{kind=link}
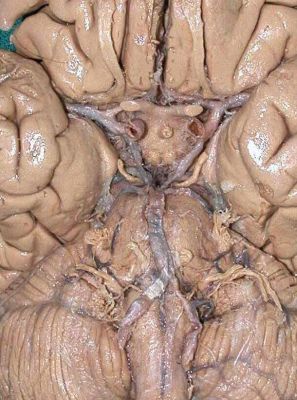{kind=link}
{kind=link}
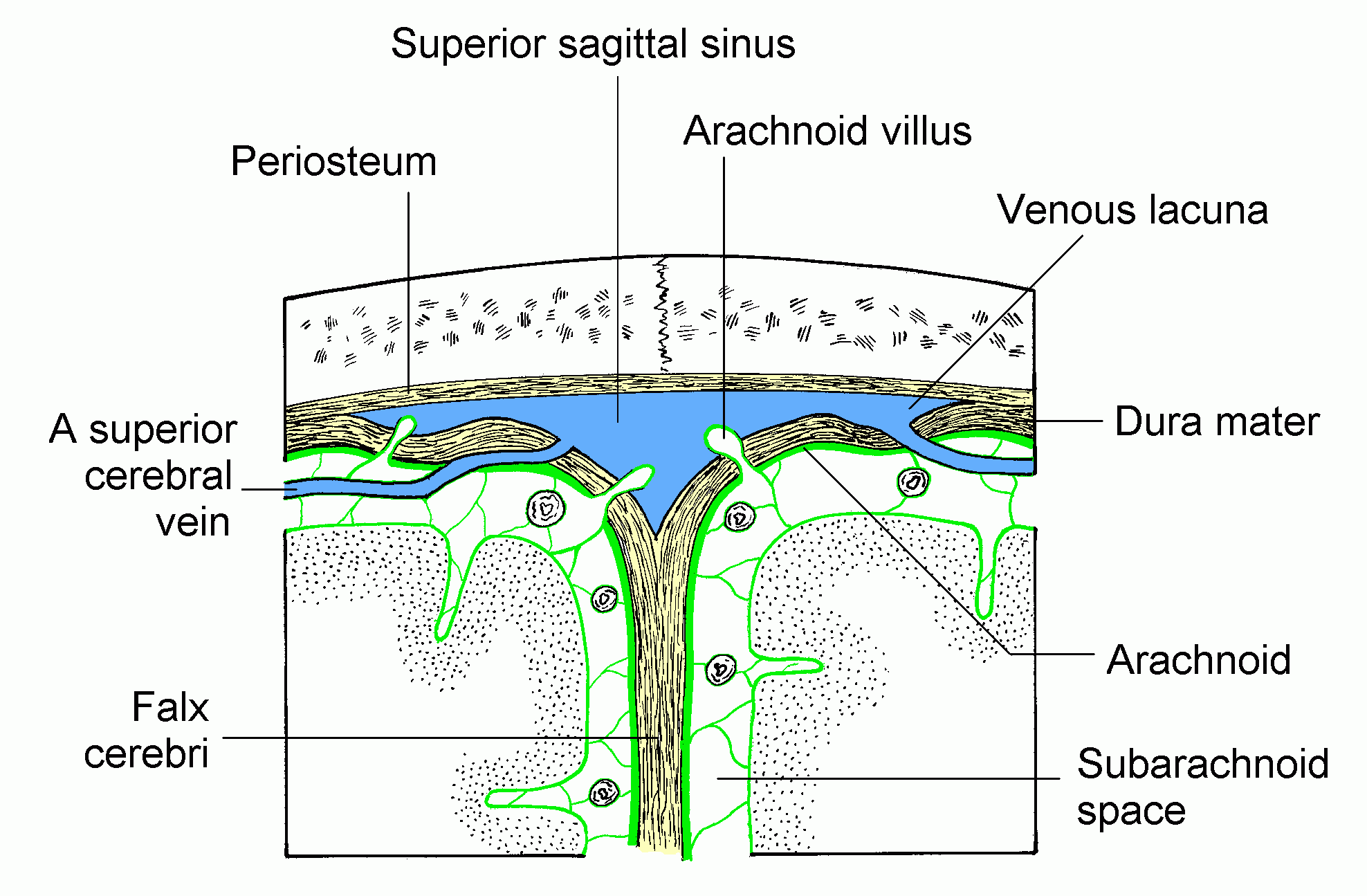{kind=link}
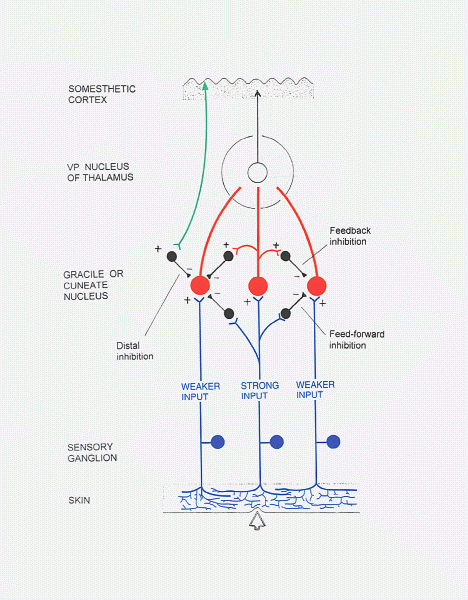{kind=link}
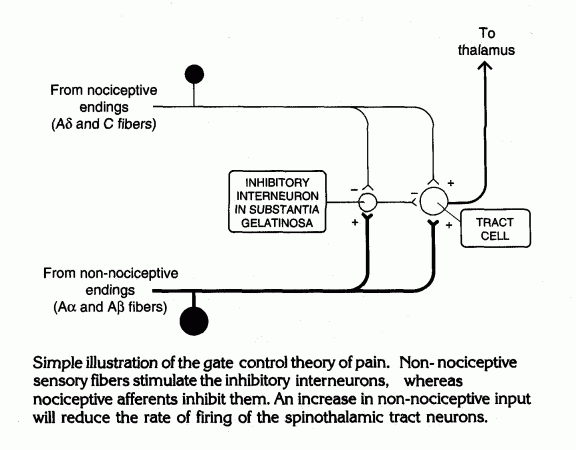{kind=link}
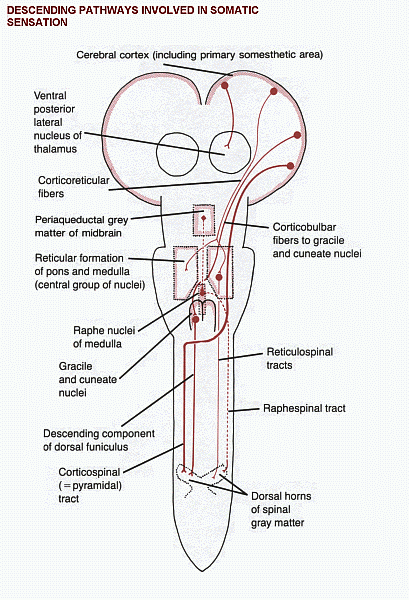{kind=link}
{kind=link}
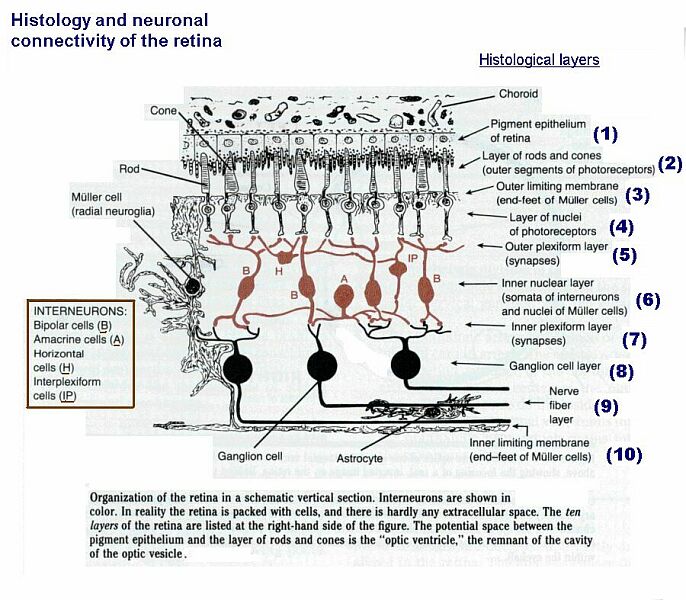{kind=link}
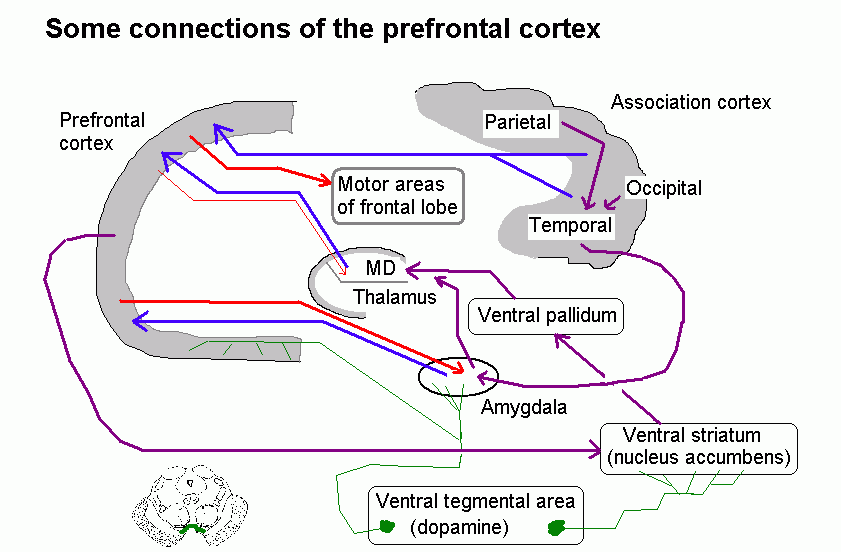{kind=link}
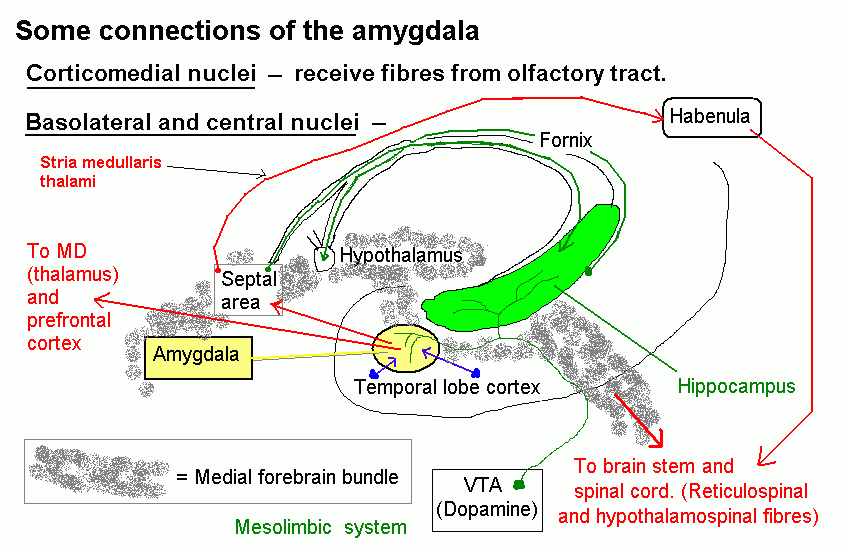{kind=link}
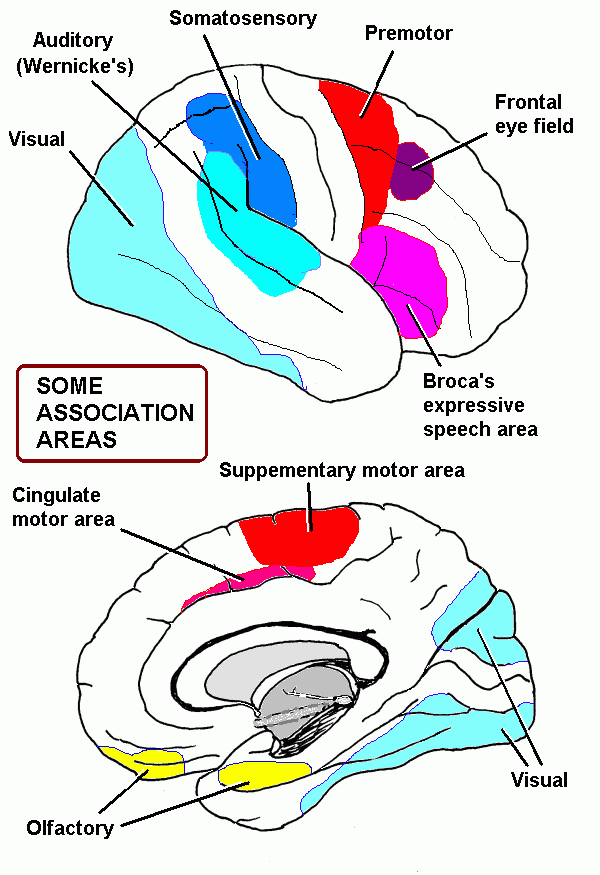{kind=link}
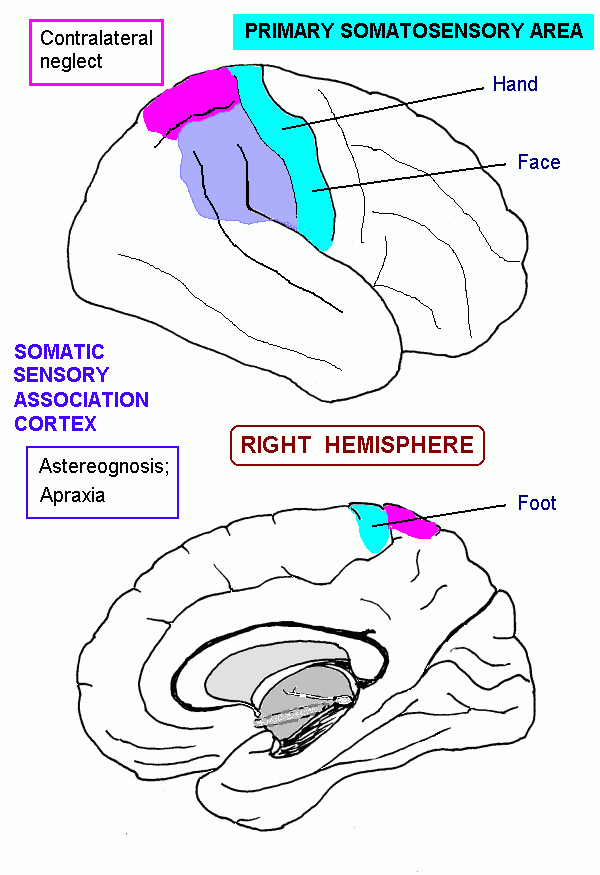{kind=link}
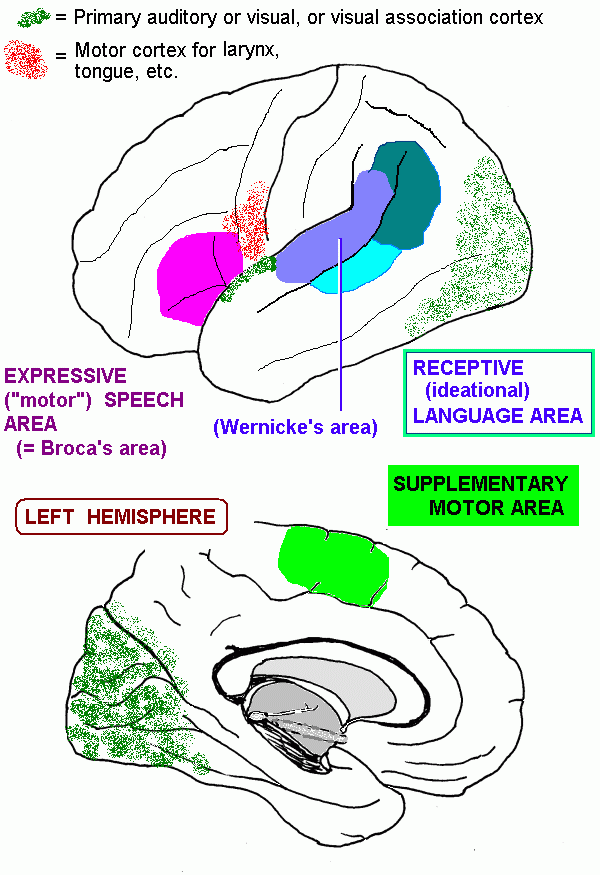{kind=link}
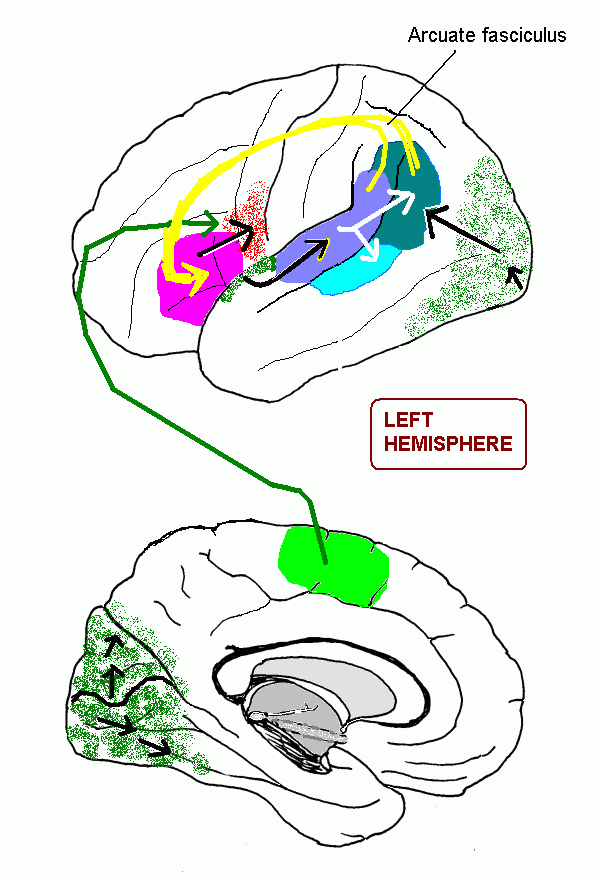{kind=link}
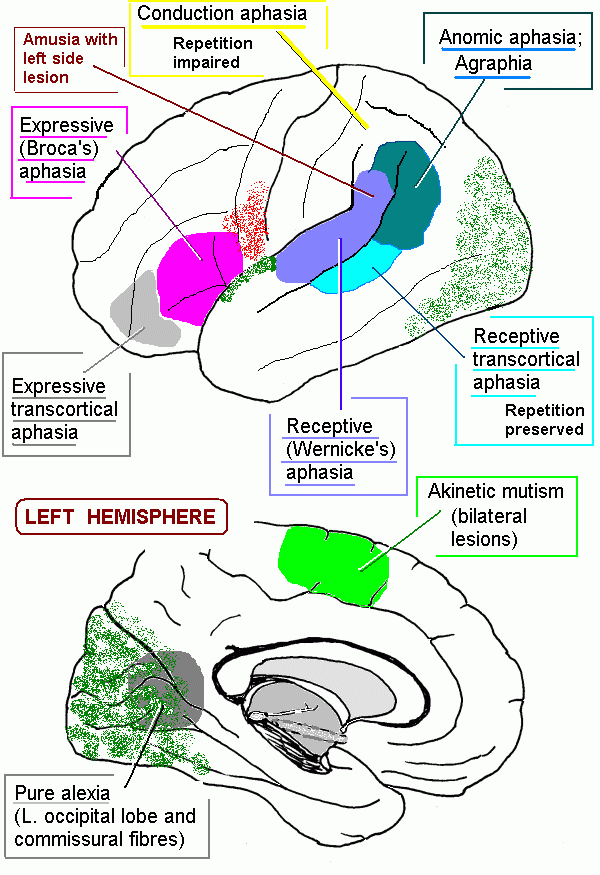{kind=link}
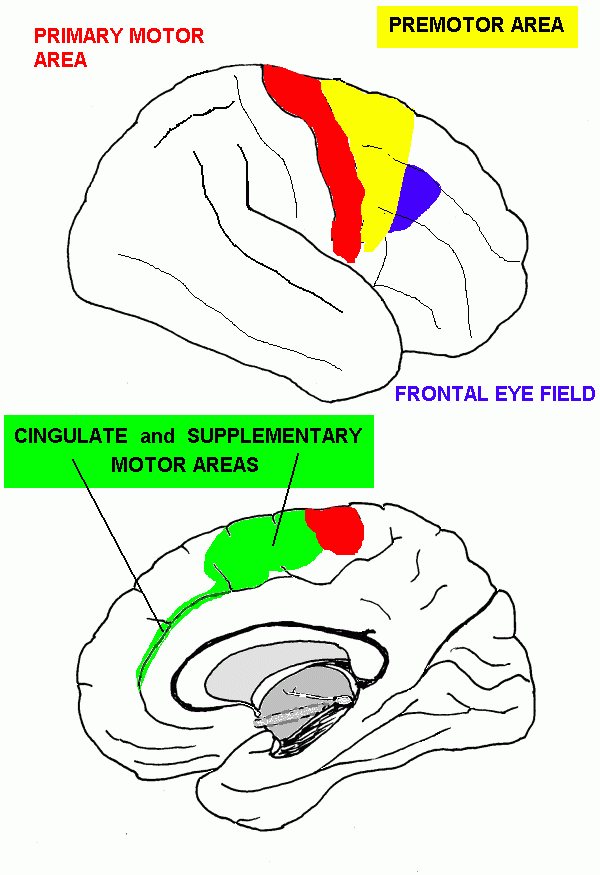{kind=link}
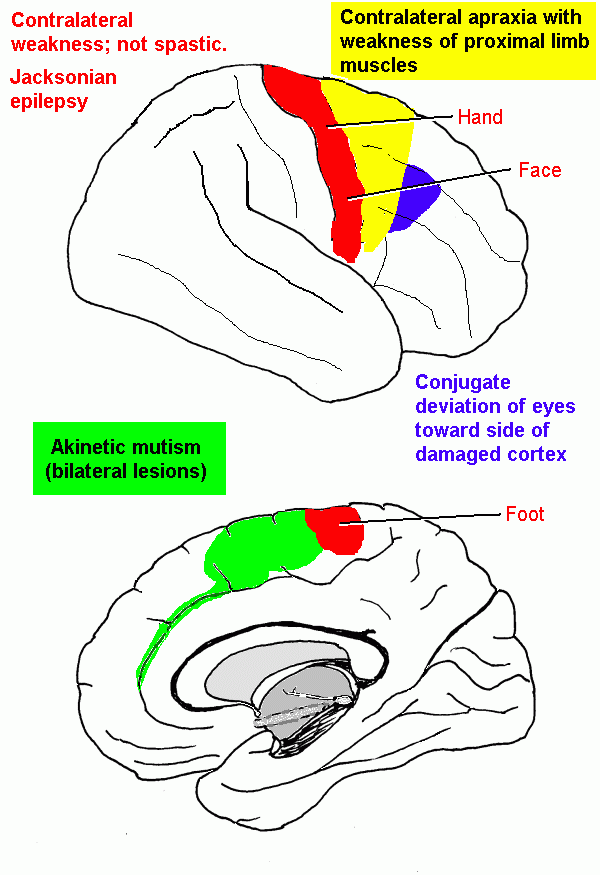{kind=link}
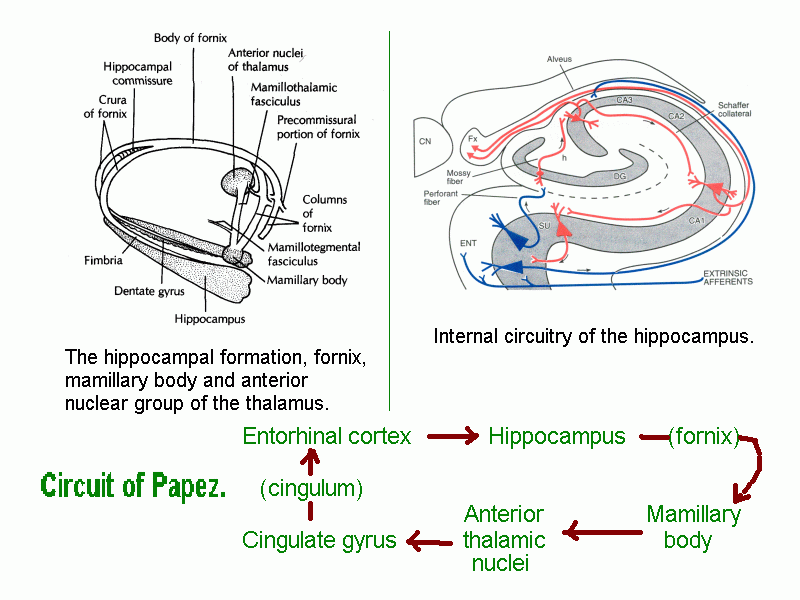{kind=link}
{kind=link}
{kind=link}